18101298214
公司首页
实验手册
小分子化合物
细胞培养和检测
定制服务
文献资料
联系我们
订购指南
公司首页
实验手册
小分子化合物
细胞培养和检测
定制服务
文献资料
联系我们
订购指南
首页
实验手册
揭示UHRF1调控有丝分裂纺锤体结构和染色体行为的新机制
发布日期:2023/10/9 18:58:00
核蛋白
UHRF1
(ubiquitin-like with PHD and ring finger domains 1)是一个具有五个结构域的表观遗传调控因子,其通过与结构域结合的蛋白协同作用维持DNA 甲基化以及组蛋白修饰。此外,UHRF1调控细胞周期转换与细胞增殖,在增殖活跃的细胞,包括多种癌症组织和细胞中异常高表达,作为一个癌基因促进肿瘤发展。反之,下调UHRF1提高了本在肿瘤细胞中沉默的p53/p21
Cip1/WAF1
表达,因而激活了DNA损伤反应,使细胞周期阻滞在G1/S期 [1,2]。此外,另一个报道显示UHRF1下调激活DNA损伤反应、导致G2/M期阻滞、诱导Caspase 8依赖的细胞凋亡 [3] 。然而,
UHRF1
在
G2/M
细胞周期转换中扮演的角色,特别是调控有丝分裂进程的具体机制仍未揭示。
该研究首次揭示了UHRF1作为核蛋白,在有丝分裂期在细胞核外通过催化驱动蛋白EG5的 K1034位点多泛素化,进而调控双极纺锤体组装和染色体结构的新功能,为UHRF1下调导致的G2/M细胞周期阻滞从机制上提供了理论证据。
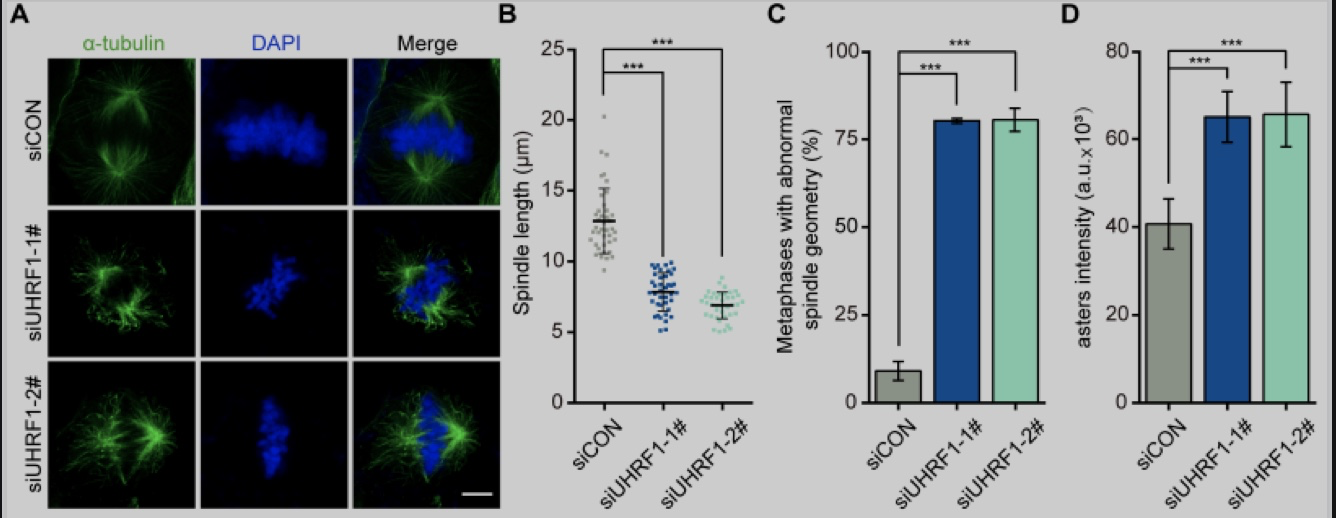
研究人员首先发现敲低UHRF1导致了细胞有丝分裂阻滞,同时这些细胞的染色体在中期赤道板上呈现出异常的不对称排列,提示UHRF1可能参与调控细胞有丝分裂正常进程,特别是组装双极纺锤体和维持染色体结构,从而保证基因组的稳定性。通过细胞免疫荧光,研究人员观察到UHRF1敲低后,
中期纺锤体的长度明显缩短,纺锤体中部的微管束向外部呈放射状分散或扭曲,同时星状体的荧光信号强度也显著增强,在UHRF1敲低的细胞中过表达突变体UHRF1
ΔRING
也获得了相同表型。
图1. UHRF1细胞有丝分裂阻滞的影响
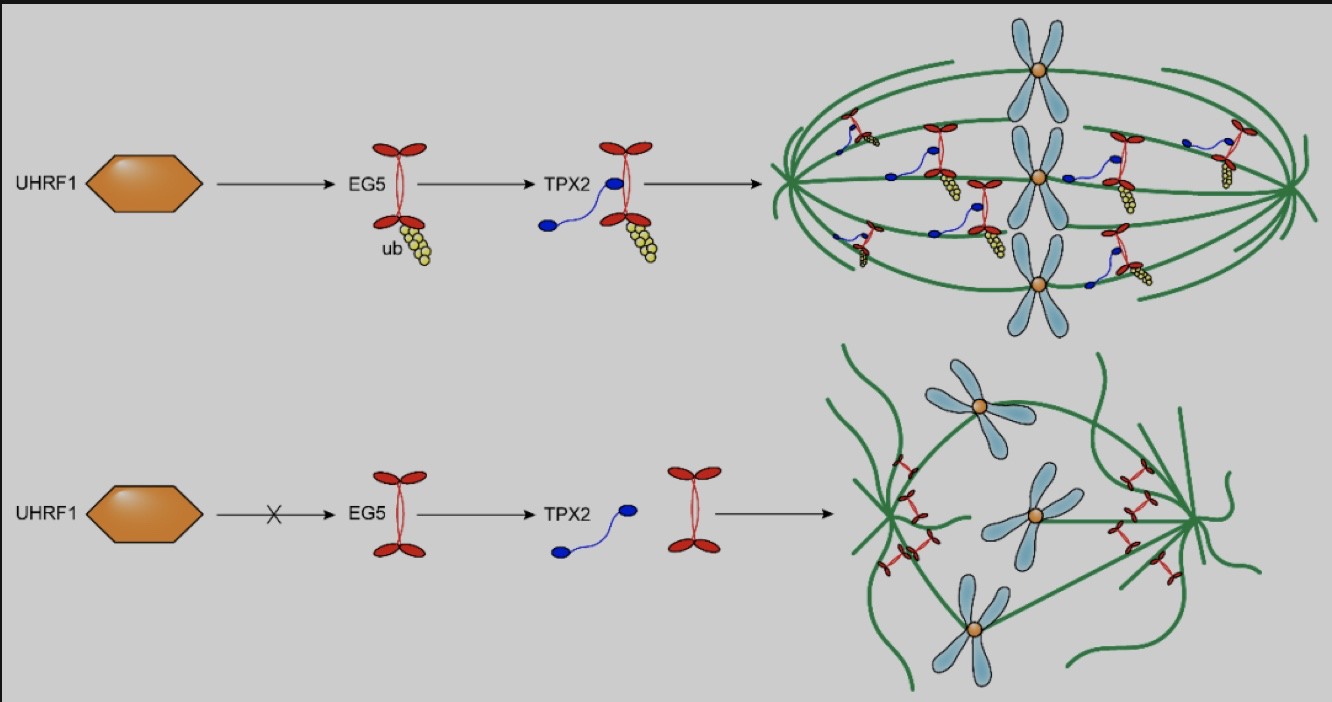
随后利用质谱、免疫共沉淀和GST-pull down实验,研究人员发现并验证了UHRF1的RING结构域与EG5的C-端tail结构域在有丝分裂期可发生直接相互作用。进一步深入探究了蛋白质互作后的生物学功能,结果表明造成该细胞表型的原因是UHRF1诱导EG5在K1034位点发生了K63多泛素化修饰,该修饰进一步促进了EG5与纺锤体装配因子TPX2的相互作用,从而确保有丝分裂中期EG5在纺锤体上的精确分布。因此,当在EG5稳转敲低的DU145细胞中分别过表达EG5
WT
和EG5
K1034R
后,相较于EG5
WT
,过表达EG5
K1034R
细胞周期仍阻滞于有丝分裂期,同时观察到明显的纺锤体缺陷和染色体排列不均,
证明UHRF1的确通过对EG5的K1034位点泛素化修饰调控了细胞周期、染色体在中期赤道板上排列,以及维持了双极纺锤体结构。
图2. UHRF1通过对EG5的K1034位点泛素化修饰调控细胞周期、染色体在中期赤道板上排列
文章结论与讨论,启发与展望
此项研究首次阐明了UHRF1作为一个核蛋白,在有丝分裂期通过催化EG5蛋白多泛素化,从而参与双极纺锤体组装和染色质结构的新功能。这种功能是完全独立于其在细胞核内行使表观遗传调控和DNA损伤修复的作用。该研究揭示了UHRF1调控有丝分裂纺锤体结构和染色体行为的新机制,并为UHRF1作为潜在的抗癌药物靶点提供了理论证据。
上一篇:
发现肝脏维持糖脂代谢平衡的新机制
下一篇:
绘制骨腱界面发育细胞图谱
已经到最底了
技术支持:
库价化学
Copyright © 2024北京螽斯羽生物有限公司 备案号:
京ICP备2023018288号-1