18101298214
公司首页
实验手册
小分子化合物
细胞培养和检测
定制服务
文献资料
联系我们
订购指南
公司首页
实验手册
小分子化合物
细胞培养和检测
定制服务
文献资料
联系我们
订购指南
首页
实验手册
Cell | 破解百年难题!清华大学最新研究
发布日期:2023/10/1 13:26:00
重力控制着植物的定向生长,一个多世纪前提出的经典淀粉-statolith假说认为,淀粉质体沉积在特化细胞中启动了重力感应,但其分子机制尚未明确。
LAZY蛋白被认为是向地性的关键调节因子,而LAZY突变体表现出明显的向地性缺陷。
该研究报道了
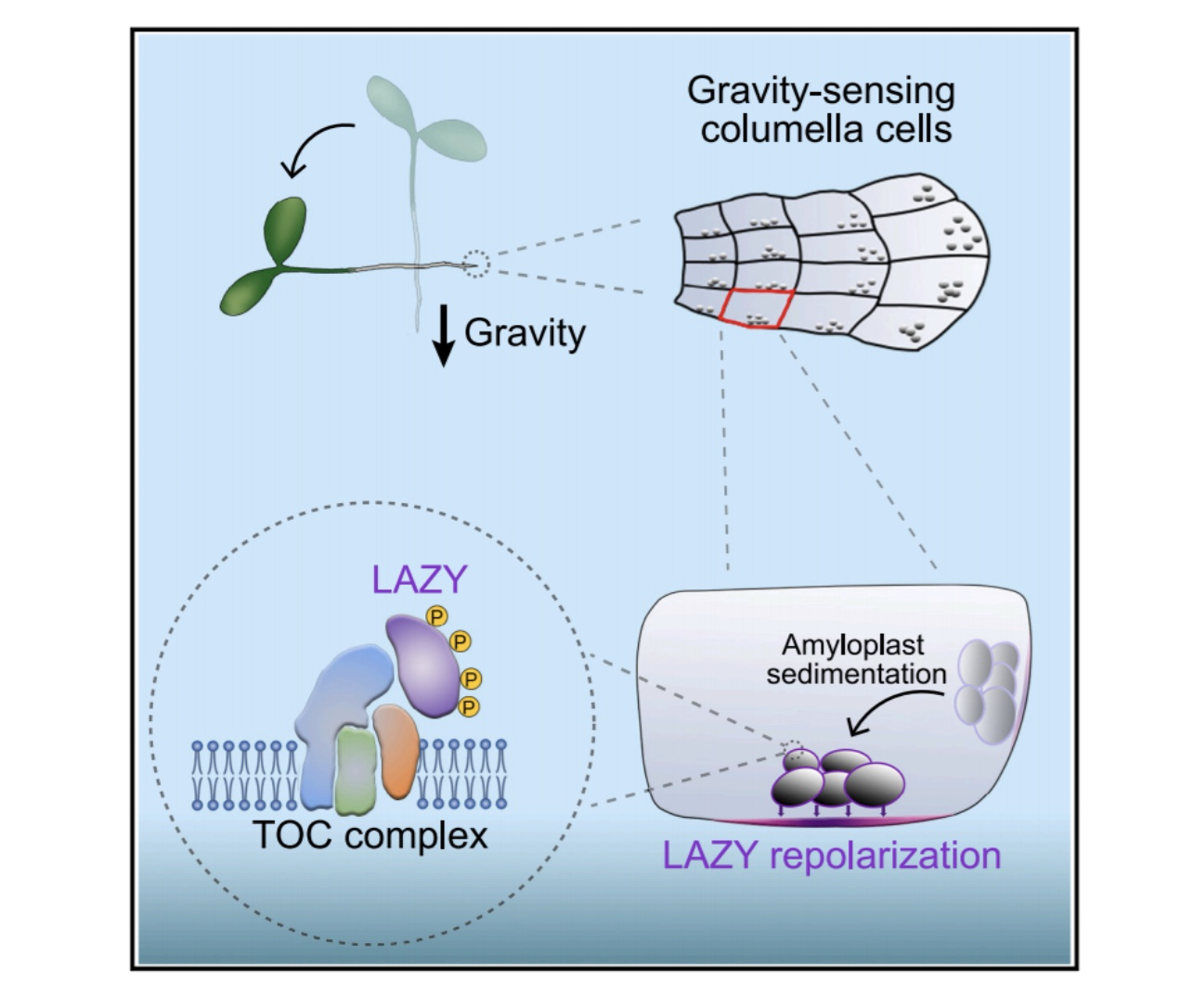
重定向的重力刺激触发丝裂原活化蛋白激酶(MAPK)信号介导的拟南芥根小柱细胞中基本极化的LAZY蛋白磷酸化。
LAZY的磷酸化增加了其与淀粉体表面叶绿体外膜(TOC)蛋白的几个转座子的相互作用,促进了淀粉体上LAZY蛋白的富集。淀粉质体沉积随后引导LAZY迁移到小柱细胞的质膜下部,在那里LAZY诱导生长素不对称分布和根的差异生长。
总之,这项研究为淀粉-statolith假说提供了分子解释:细胞器运动触发分子极性形成。
重力是影响地球上所有生物的关键环境因素
。
在植物中,重力通常引导根系向下生长(正向地性),引导地上部分向上生长(负向地性),从而控制抗旱性、养分吸收等农业性状。重力向性包括三个关键过程:重力传感、信号转导和差异生长响应。
由于地球表面上重力的方向和大小几乎是恒定的,因此,重力倾向被认为是一种姿态控制,通过感知器官相对于重力矢量方向的倾斜来触发。
淀粉-statolith假说和Cholodny-Went理论分别是解释重力感应和响应的教条
。
淀粉-statolith假说是在120年前提出的,其中认为静止细胞中淀粉体(淀粉填充的质体)的沉积是重力感应的开始。
根的小柱细胞和茎的内胚层细胞被证明是感知重力的静态细胞,遗传破坏或激光消融导致这些细胞完全破坏倾斜性
。
重力传感又分为感知(物理响应)和从物理信息到生理信息的信号转换。磷酸葡萄糖糖化酶1 (PGM1)突变体的无淀粉质体沉积受损,导致根和芽的向地性反应减慢。相反,肌动蛋白的破坏加速了淀粉体沉积,从而促进了向地性。一般认为,淀粉体沉降是重力感应的感知步骤,但淀粉体沉降的分子作用和信号转换的潜在机制尚不清楚。
Cholodny-Went理论提出,生长弯曲是由于植物弯曲器官两侧生长素分布不均,生长素外流转运蛋白PIN蛋白后来被证明有助于生长素的重定向和随后的倾斜性差异生长。
在20世纪30年代,一种不寻常的玉米突变体,其茎向地面生长,由于其匍匐表型,被描述为“lazy”。
随后又鉴定出具有相似表型的水稻lazy突变体。从那时起,LAZY家族基因被证明对许多植物物种的芽和根的向地性反应至关重要,包括水稻、玉米、拟南芥、苜蓿、家李和日本莲花。高阶lazy突变体在拟南芥中表现出比单突变体更夸张的表型,尽管不同群体报告的表型存在差异。光通过控制LAZY4的表达水平来调节向地性。最近的一项研究表明,拟南芥LAZY4 (LZY3)蛋白在侧根小柱细胞的质膜上呈极性分布,并从细胞质向质膜募集RCC1样结构域(RLD)蛋白,促进PIN3的再定位,调节生长素的流动。
LAZY4 (AtDRO1)也有报道显示核定位,但其在那里的作用尚不清楚。
文章模式图(图源自
Cell
)
Altered Response to Gravity(ARG1)编码 DnaJ样蛋白,其突变延迟了根和下胚轴的向地性
。
尽管toc132
Q730Stop
、toc75
G658R
或arg1单突变体表现出弱或无向地性表型,但叶绿体(TOC)外包膜Translocons的组分TOC34、TOC75、TOC120或TOC132的突变显著增强了arg1的向地性缺陷。
TOC蛋白的经典功能是将蛋白质导入质体,但TOC蛋白是否通过将未知信号因子导入淀粉质体来调节向斜性尚不清楚。
丝裂原活化蛋白激酶(MAPK)级联在真核细胞中高度保守
。
在植物中,MAPK信号模块调节生长和环境反应的许多方面。MKK4/MKK5-MPK3/MPK6模块调控气孔发育和定形、花序结构、侧根发育等。
MKK7-MPK6模块正调控下胚轴倾斜。
该研究报道了重力刺激触发MKK5-MPK3模块磷酸化LAZY蛋白,这可能通过与TOC蛋白相互作用促进LAZY蛋白在淀粉质体表面富集
。
然后,淀粉体沉积促进了小柱细胞中LAZY蛋白向新的质膜下侧的易位,其中LAZY蛋白从淀粉体表面重新定位到邻近的质膜是一个重要的过程。LAZY蛋白的复极化导致生长素分布不对称,最终导致生长差异。
因此,该研究揭示了淀粉体沉积的分子作用,为淀粉-statolith假说的分子机制提供了见解。
上一篇:
跌下神坛!生酮饮食或是一种促进疾病的饮食,增加心脏病、癌症风险
下一篇:
合并感染甲型流感病毒可增强SARS-CoV-2的传染性
已经到最底了
技术支持:
库价化学
Copyright © 2024北京螽斯羽生物有限公司 备案号:
京ICP备2023018288号-1