固有淋巴细胞(Innate Lymphoid Cell,ILC)是一类缺乏抗原特异性受体的淋巴细胞,主要包括ILC1,ILC2,ILC3三个亚群,其分化谱系与T、B淋巴细胞一样,来自共有淋巴祖细胞(CLP),可由淋巴祖细胞样细胞(αLP)、早期淋巴前体细胞样细胞(EILP)、共有辅助淋巴前体细胞样细胞(CHILP)、CLP前体细胞(ILCP)等发育分化为成熟的ILC细胞。
ILC的生物学意义:ILC细胞能够通过与周围环境中的神经细胞、上皮细胞、基质细胞、适应性免疫细胞、髓系细胞、共生菌群等相互作用,协调环境中的信号并广泛参与抗病原体感染、炎症疾病发生、器官形成及组织修复、癌症发生、代谢及生物节律等生物学过程。
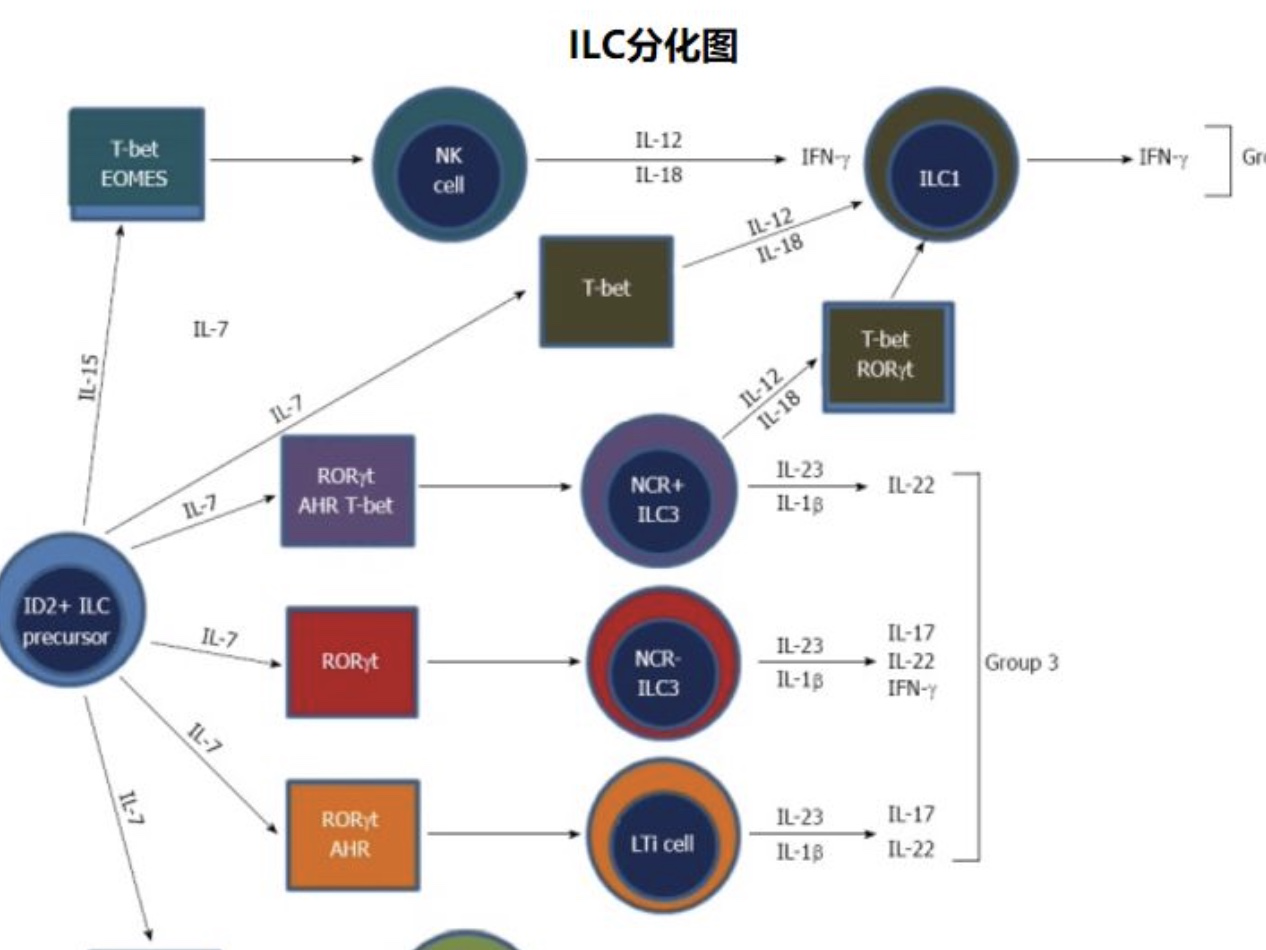
固有免疫细胞均来源于ID2阳性祖细胞。
ILC1产生IFN-γ;ILC1的分化发育机制尚未完全阐明,已知其发育所需的转录因子为t-bet。
ILC2产生IL-5和IL-13;ILC2的分化依赖于转录因子GATA3和ROR。
ILC3产生IL-17、IL-22和IFN-γ。ILC3的分化依赖于RORT。此外,ILC3的亚群需要额外的转录因子,如芳基烃受体(AHR)来分化。
NK细胞需要IL-15进行刺激,其他ILC需要IL-17来刺激。NK细胞在t-bet和中胚层上皮素(eomes)作用下可分化为ILC1。
(Reference:Immunopathology of inflammatory bowel disease)
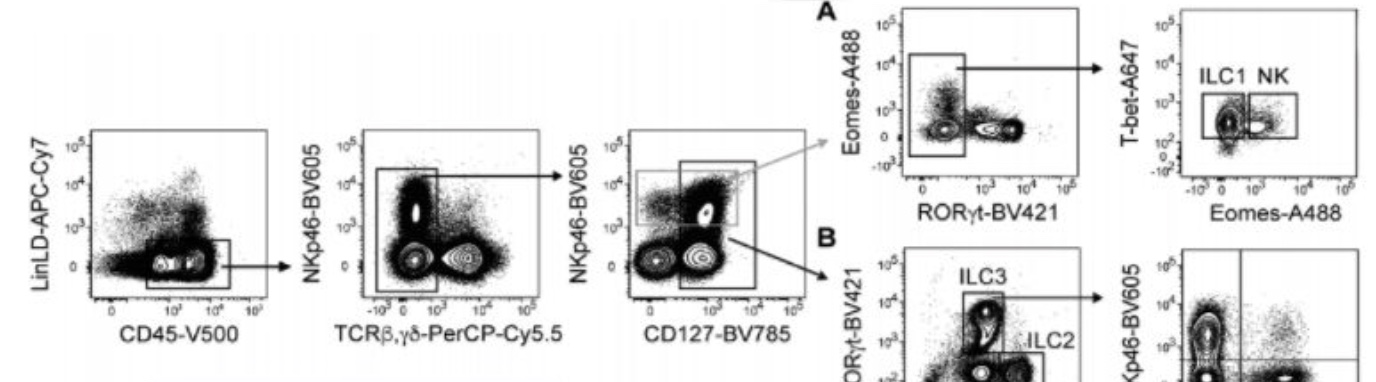
Mouse固有免疫细胞分型流式检测
Human固有免疫细胞分型流式检测

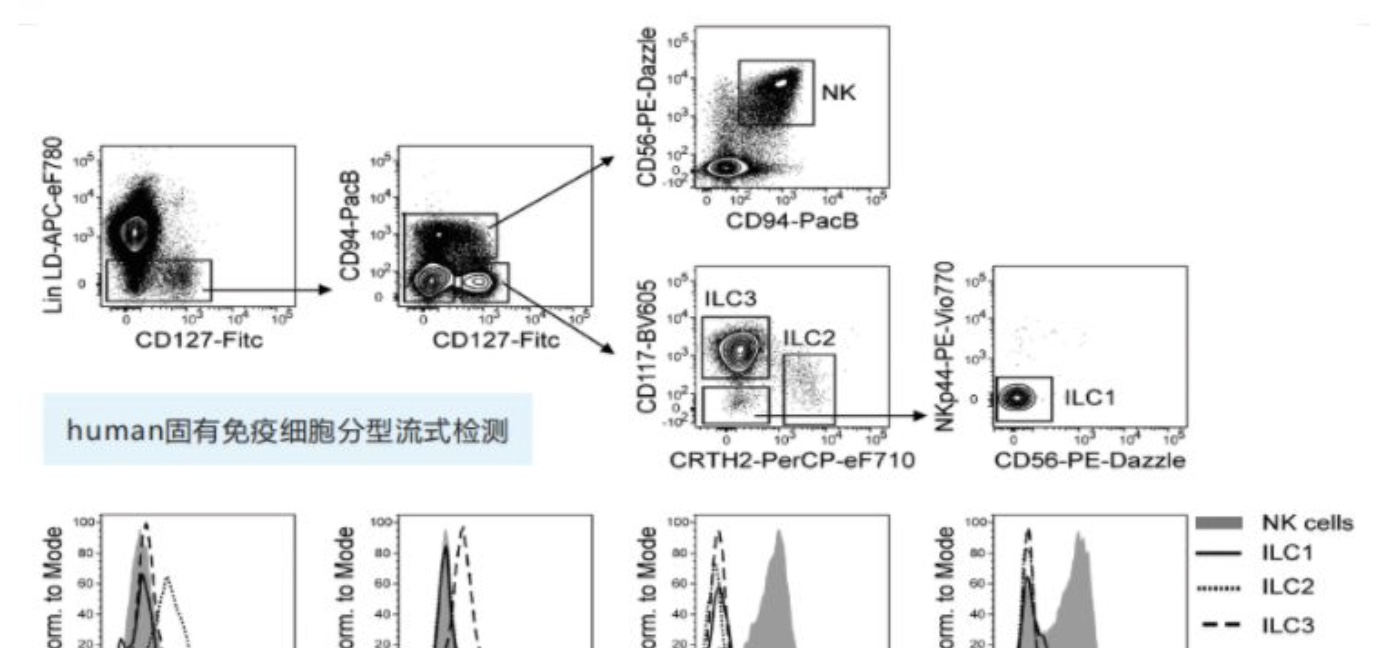
(Reference: Guidelines for the use of flow cytometry and cell sorting in immunological studies)
ILC1: 表达T-bet和IFN-γ,有助于1型炎症
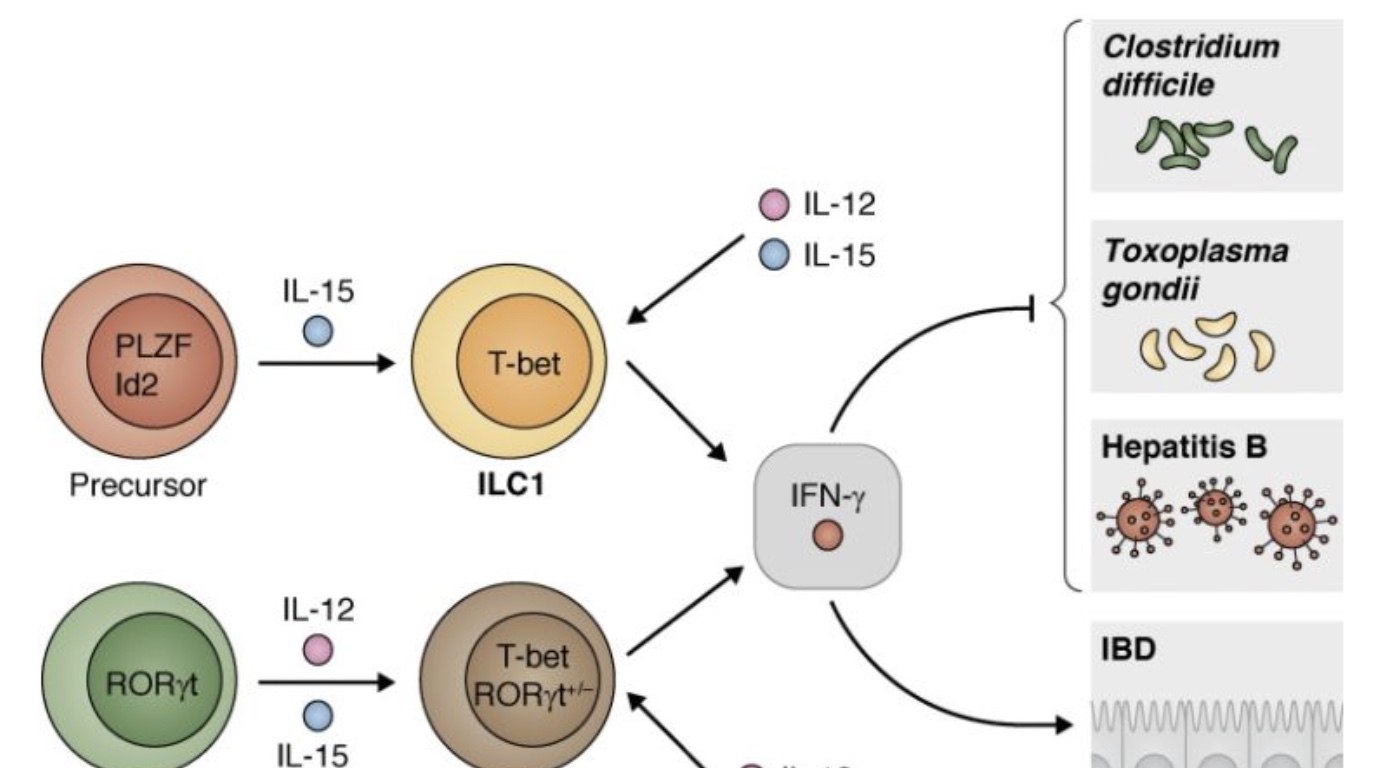
表达T-bet的ILC1s由表达PLZF和ID2的祖细胞发展而来,IL-15可刺激其分泌细胞因子。表达RORγT的细胞可以失去RORγT表达,对IL-12信号的反应中获得T-bet表达。IL-12和IL-15对表达有T-betRORγT的ILC或T-bet+ILC1s有刺激作用,产生在艰难梭菌、弓形虫和乙型肝炎等病原体感染期间具有保护作用的IFN-γ。表达T-bet的ILC所表达的RORγT在感染期间特别重要,尽管这些ILC在类型上很像ILC3,但在肠易激综合征的情况下,它们也可以导致肠道炎症和病理性病变。(Reference: Emerging concepts and future challenges in innate lymphoid cell biology)
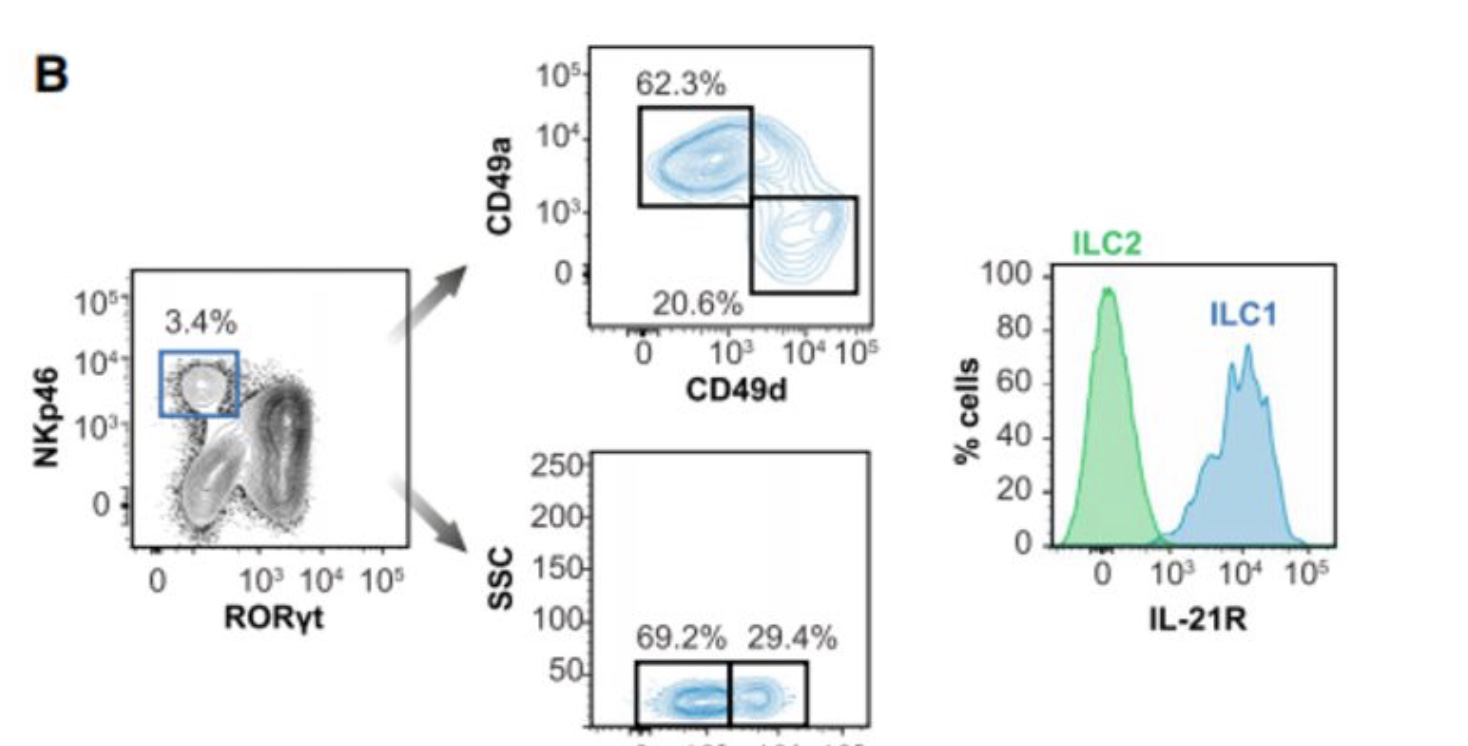
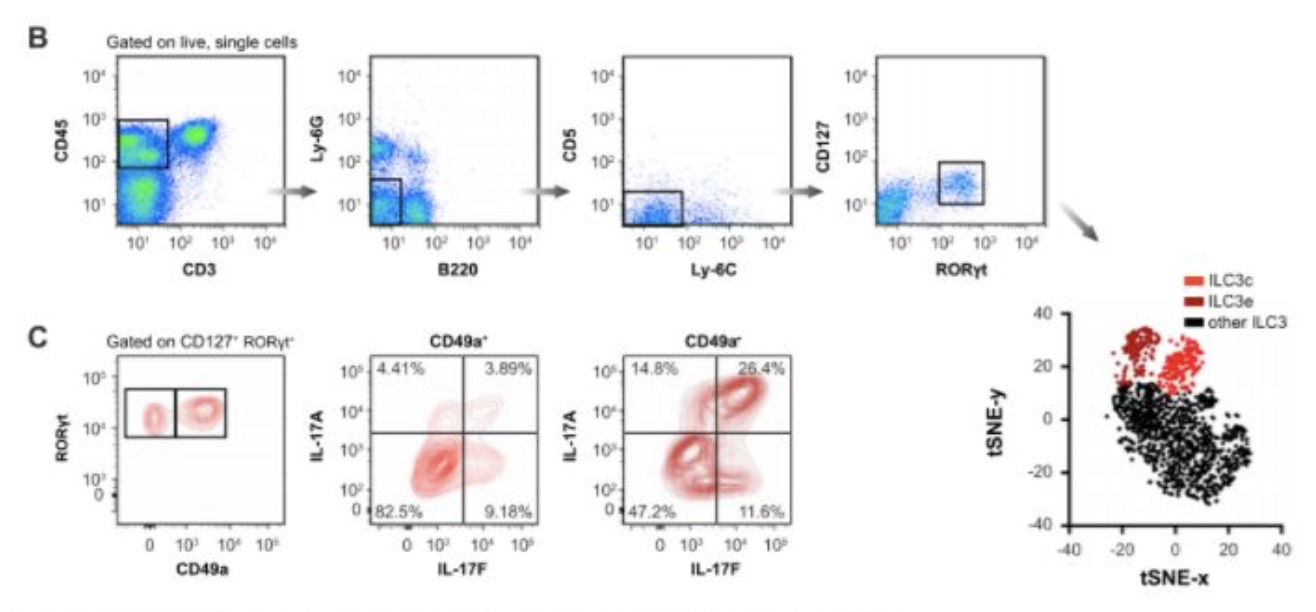
流式细胞术分析证实小肠RORγT-NKp46+群体中存在ILC1亚群,可根据其表达的CD49a(itga4编码)以及CCL5产生细胞的子集进行分群。
ILC2:在健康和疾病中的激活、调节和作用
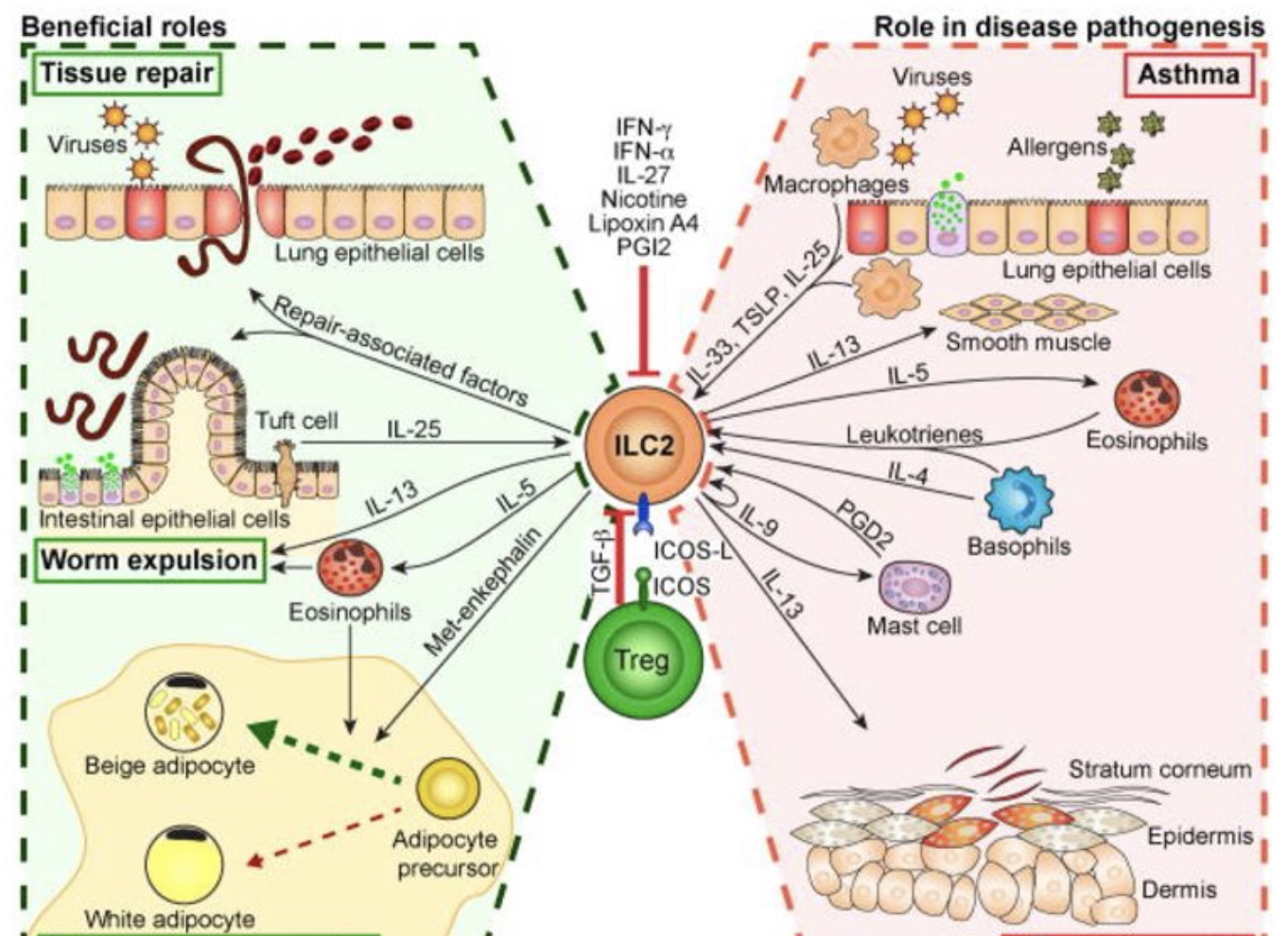
ILC2可以被多种刺激物激活,如细胞因子IL-33、TSLP、IL-25、类花生酸、PGD2和白三烯。ILC2通过产生IL-5和IL-13在哮喘和特异性皮炎的发病机制中发挥重要作用。ILC2可以通过IL-13修复蠕虫迁移或病毒感染引起的粘膜组织损伤以及修复相关基因。ILC2还可以通过将白色脂肪细胞转化为米色脂肪细胞来控制肥胖。尼古丁、干扰素、脂蛋白A和PGI2以及调节T细胞(Treg)通过ICOS/ICOS-L相互作用和TGF-β抑制ILC2。

流式细胞术分析证实ILC存在产生IL-4的亚组,其不同于产生IL-5的亚组。IL-5产生与KLRG-1和Sca-1的高表达相关,而IL-4产生在KLRG-1细胞中最高。
ILC3:在维持肠道内环境平衡和炎症性肠病发生中的应用
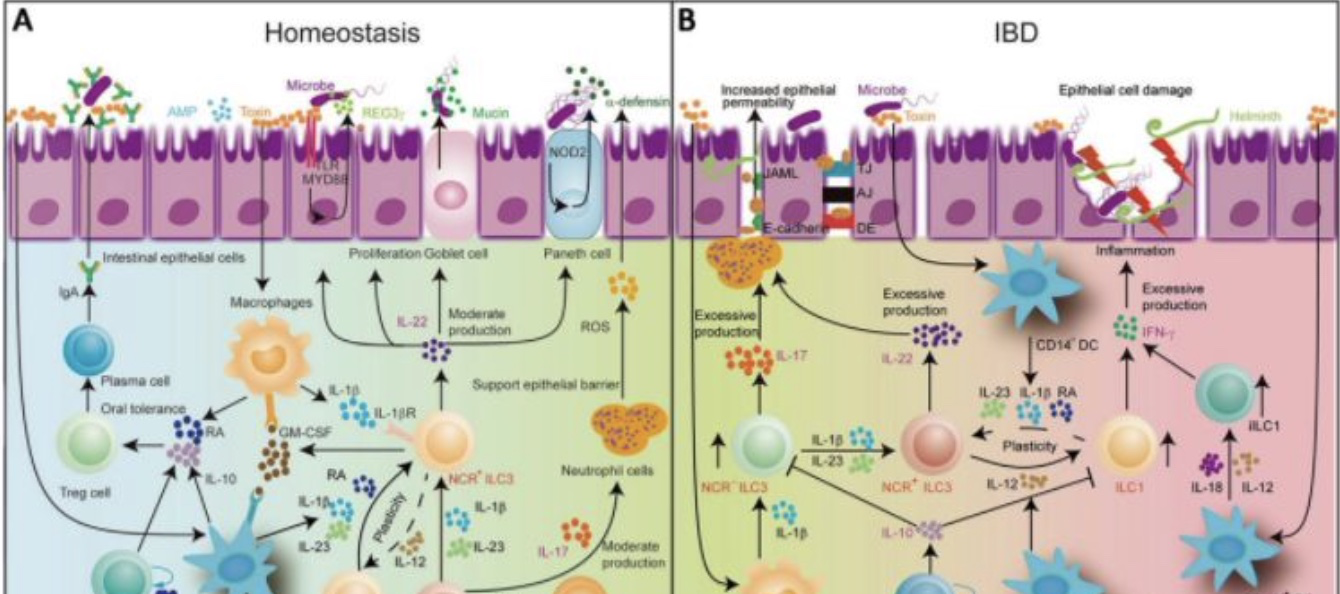
巨噬细胞被细菌刺激后产生IL-1β,IL-1β与ILC3上的IL-1受体结合(IL-1R)结合,促进IL-22、IL-17和GM-CSF的释放。GM-CSF触发树突细胞(DC)和巨噬细胞生成维甲酸和IL-10,进而促进Treg形成。IL-22促进上皮屏障完整性和增殖,诱导AMPS、REG3γ和黏连蛋白的产生。IL-17可以刺激中性粒细胞,同时支持上皮屏障保护。表达MHC II的ILC3可以抑制共栖特异性CD4+T细胞。NCR-ILC3可通过IL-1β与IL-23刺激转换为NCR+ILC3。(Reference: ILC3 function as a double-edged sword in inflammatory bowel diseases)
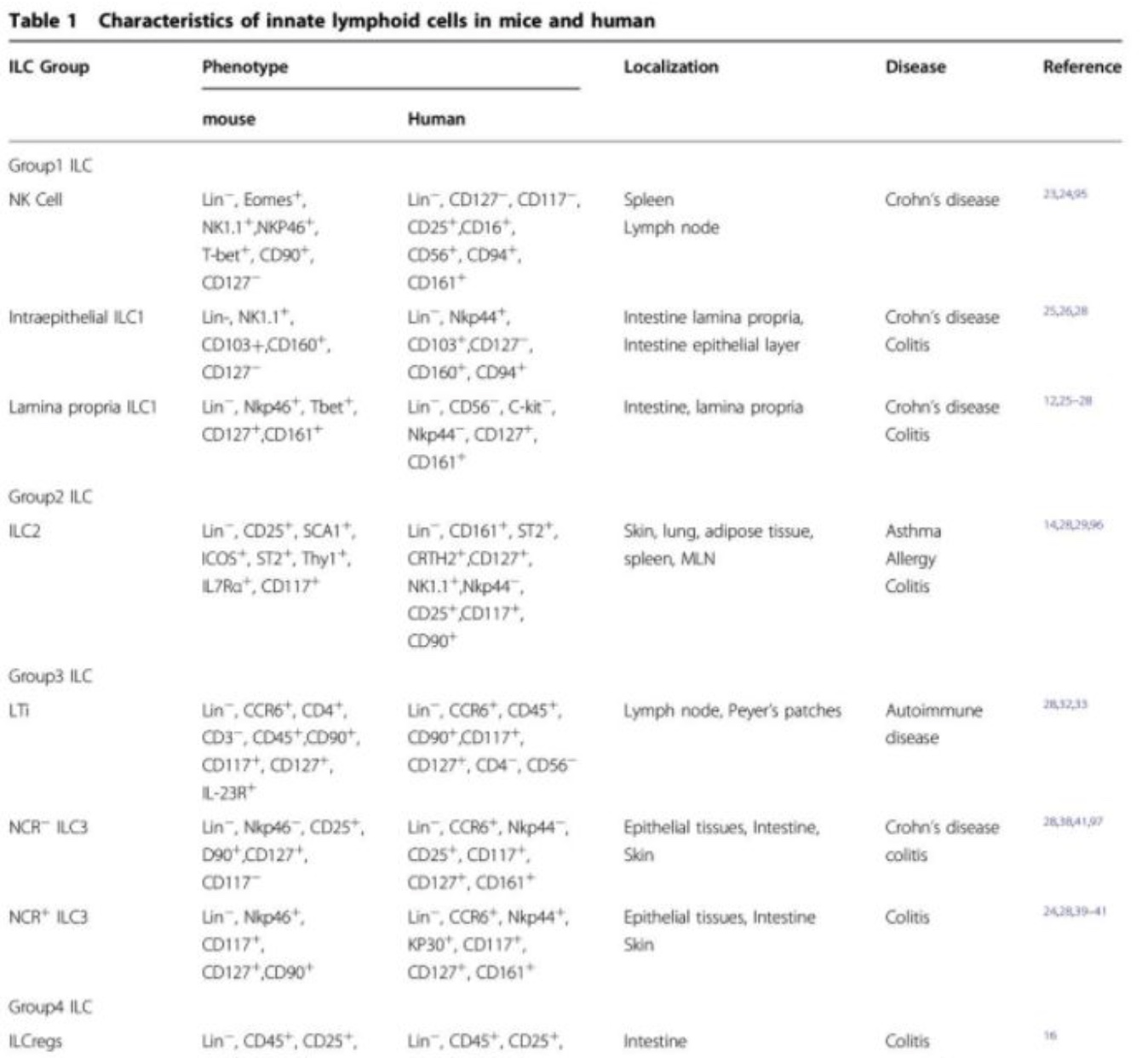
Characteristics of ILC in mice and human