
PIWI蛋白相互作用RNA(PIWI-interacting RNAs,piRNAs)对动物生育能力至关重要。PIWI蛋白是生殖细胞中表达的Argonaute蛋白的一个特殊家族。piRNAs在所有两侧对称动物中的保守功能是沉默序列互补的转座子(transposable elements,TEs)[1-4]。成年哺乳动物表达高水平粗线期piRNAs,以极快的速度进化,其与果蝇和斑马鱼中发现的富含TE的原始piRNAs或哺乳动物前粗线期piRNAs不同的两个特征是:(1)粗线期piRNAs在减数分裂的粗线期表达;(2)绝大部分并不源自TE序列。虽然大多数粗线期piRNA缺乏明显的靶标,但粗线期piRNA位点的拷贝数和核苷酸序列都不保守[5,6,7],其中许多piRNA位点甚至在亲缘关系密切的哺乳动物中也不存在[8,9]。粗线期piRNAs被认为可以序列特异性的调节mRNAs[10-14]或序列非特异性的稳定PIWI蛋白[15]。这种说法很难解释粗线期piRNA为何会快速进化,这种快速进化和它们在基因组上多个位点的冗余分布也使它们的功能研究变得复杂。因此,粗线期piRNAs有什么功能,是什么促进了它们的快速进化,目前还不清楚。报道了羊膜动物利用固有的遗传不稳定性来保护生殖细胞基因组的完整性新机制。研究通过对鸡、鸭、小鼠和人的比较研究,以及对不同鸡品系的长阅读纳米孔测序,发现羊膜动物的piRNA位点有以下三个机制驱动快速进化:(1)结构变异(Structural variations ,SVs)的高局部突变率;(2)开始于生殖细胞发育过程中的减数分裂粗线期抑制年轻和活跃的TEs的正选择;(3)和选择清除有害SV热点的负选择。结果表明,粗线期piRNA位点的遗传不稳定性,在产生某些致病性SVs的同时,也通过驱动快速进化的piRNA序列形成来保护基因组完整性免受TE的插入。

鸟类粗线期piRNAs迅速进化。为了了解粗线期piRNAs是否是哺乳动物所特有的。研究者收集来自鸡八个关键发育阶段的睾丸来研究鸡piRNA在精子发生过程中的动态变化特点(图1a,i)。大多数piRNAs在从12周到18周间发生的的第一次减数分裂时期表达(图1a, i和iii),这一阶段与CIWI(PIWI基因)的mRNA的表达相一致,CIWI在小鼠体内的同源基因与粗线期piRNAs特异性结合[16] (图1a, iv)。研究在粗线期精母细胞的胞质中检测到CIWI蛋白的特异性染色(图1b) 。研究绘制了八个发育阶段中每个piRNA位点的piRNA丰度,在粗线期检测到大量表达,在早期阶段几乎没有piRNA存在(图1c),表明成年鸡睾丸中的大多数piRNAs是粗线期piRNAs。与哺乳动物粗线期piRNA相似,成年鸡睾丸中的大多数piRNAs不是来自重复区域或蛋白编码基因区域(图1a, iii)。这证明鸡粗线期piRNAs的存在,表明粗线期piRNAs在鸟类和哺乳动物生殖细胞发育过程中发挥作用。为了测试鸡粗线期piRNA位点在近缘鸟类中的保守性,研究者在鸭睾丸中检测到大量的piRNAs, 但与鸡piRNA位点同源的136个位点不产生piRNAs(图1d)。与鸭比较表明,鸡piRNA位点在序列水平(是否存在同源区)和功能水平(是否同源区产生piRNAs)上都经历了快速的增加和丢失。与鸡和鸭之间mRNAs和长非编码RNAs(lnc RNAs)的外显子和内含子以及一组随机改组对照相比,piRNA位点在脊椎动物中(图1e左)和鸟类体内(图1e右)表现出最迅速的进化速度。研究结论是:快速进化是哺乳动物和鸟类粗线期piRNAs的一个共同特征。考虑到与哺乳动物基因组相比,鸟类基因组在核苷酸序列、基因合成和染色体结构方面表现出高度的进化停滞[17],哺乳动物和鸟类之间快速进化的共同特征证明存在驱动粗线期piRNAs进化的统一驱动力。
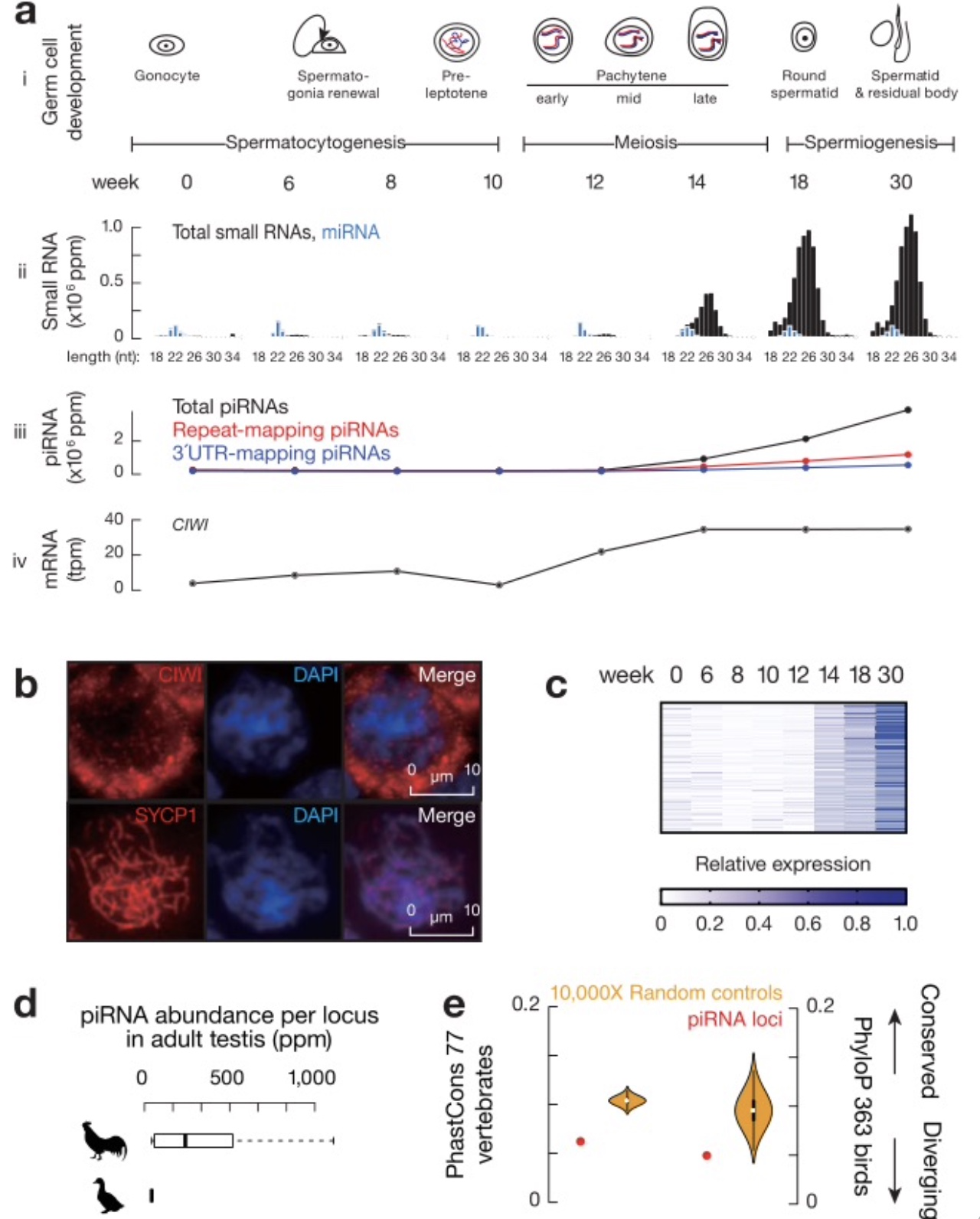
图1 鸡体内粗线期piRNAs的存在
(图源:Sun YH, et al., Nat Commun. 2023)
piRNA位点是鸟类和哺乳动物中的SV热点。研究者决定使用鸡作为模型来分析粗线期piRNA位点在较短的进化时间尺度内的突变事件。研究者对六种不同品种的鸡进行了测序,这些鸡具有不同的地理分布和特定的表型。为了高分辨和高保真地捕获结构变异(SVs,影响≥ 50 bp的突变)[18,19],研究者使用纳米孔技术ONT进行测序,并达到每只鸡31X的测序深度,与野生鸡参考基因组相比,每种家养鸡平均有17321±777个SV事件(图2a, b)。粗线期piRNA位点占鸡基因组的0.98%[20],但SVs出现在粗线期piRNA位点上的频率更高:12.4%的串联重复(189/1526),19.4%的倒位(26/134),1.7%的缺失(314/18721),以及1.2%的插入(165/13442)与piRNA位点重叠(图2a, iii)。研究发现在piRNA位点中SVs如串联重复、倒位和缺失(图2c)在piRNA位点上显著富集。鸡粗线期piRNA位点与这些SV热点显著重叠。因此鸡piRNA位点同时也是SV热点。
为了测试piRNA位点的SVs对piRNA多态性的影响,研究者对六种鸡小RNAs进行了单独测序。尽管与大规模基因组相比,插入在piRNA位点没有显著富集(图2a, c),但插入的片段带来了新的piRNA序列的产生。piRNA位点中的SVs也造成了piRNA表达丰度(图3a)、表达有义/反义方向(图3b)以及不同piRNA香浓多样性指数的显著增加(图3c)。因此,piRNAs与SV热点重叠是造成piRNAs的快速进化的第一个机制。
研究者接着想知道SV热点和piRNA位点之间的联系是鸟类特有还是所有羊膜动物共有。粗线期piRNA位点,而不是前粗线期piRNA位点,与人类SV热点显著重叠(图 3e)。说明和鸡piRNA位点一样,人粗线期piRNA位点也是SV热点。因此,SVs的高局部突变率是一种保守机制推动粗线期piRNAs在羊膜动物中的快速进化。
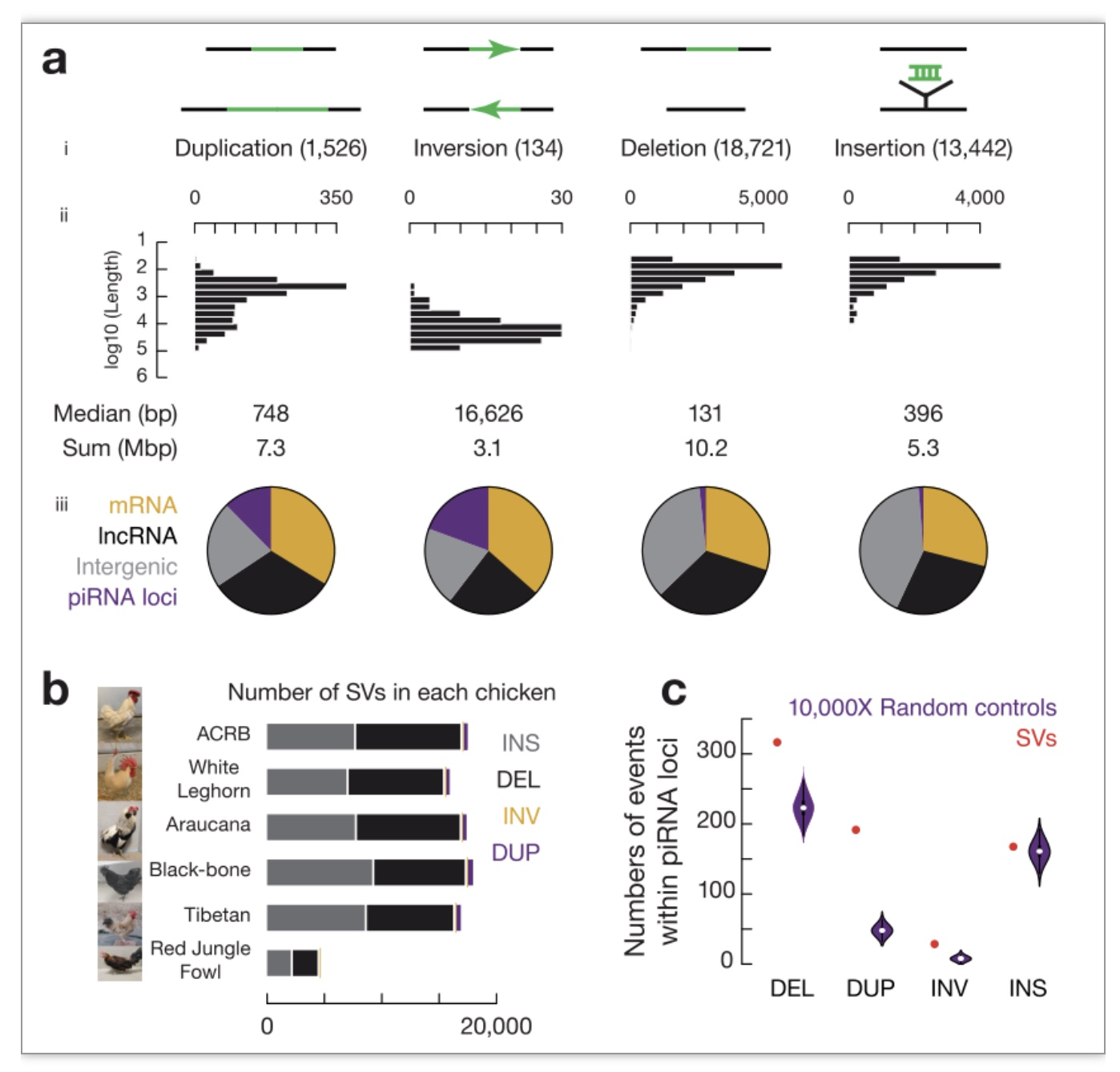
图2 鸡piRNA位点是SV热点
(图源:Sun YH, et al., Nat Commun. 2023)
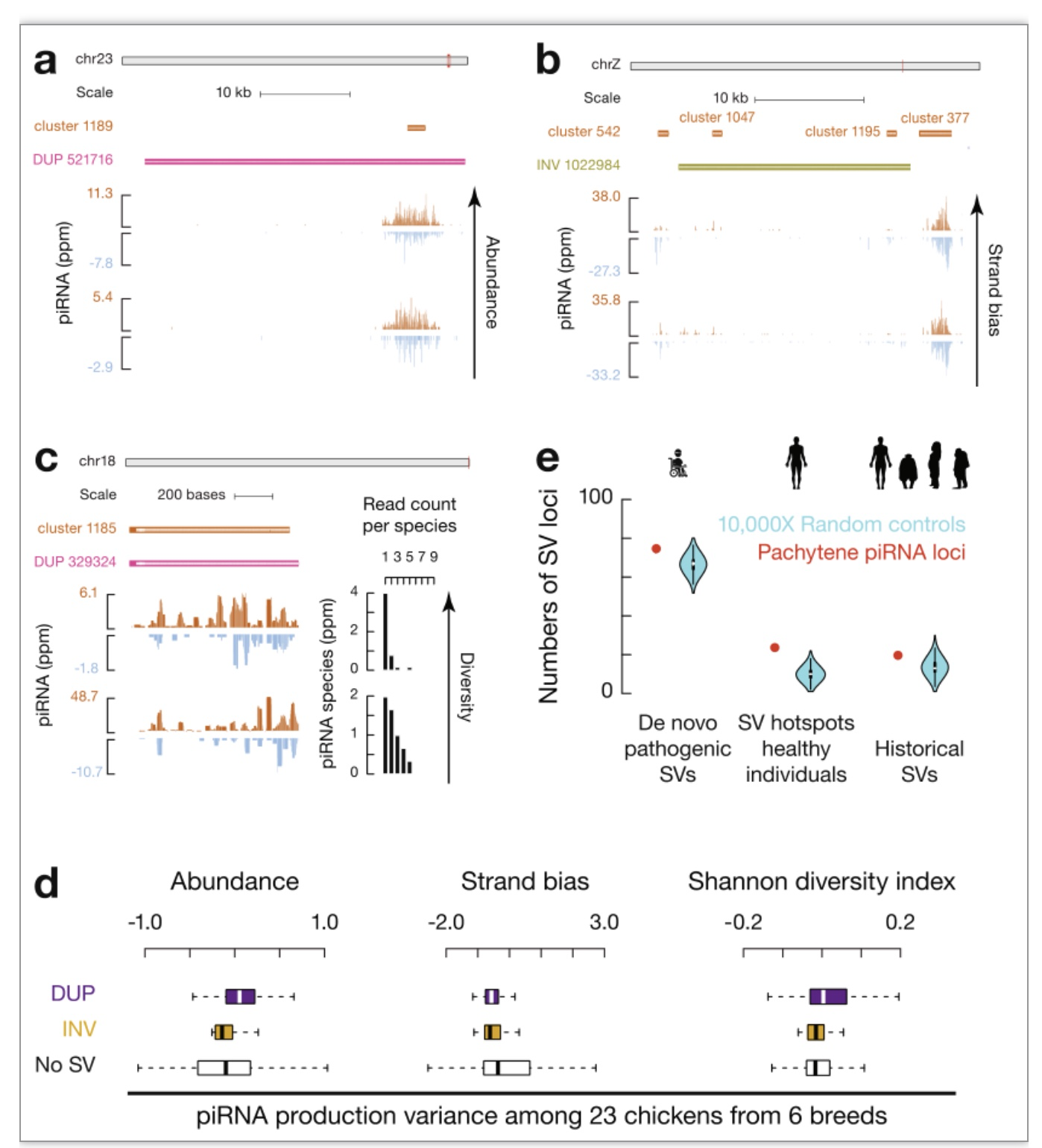
图3 保守机制以实现piRNA可塑性
(图源:Sun YH, et al., Nat Commun. 2023)
与SV热点重叠的piRNA位点的趋同进化。研究设想了三种可能的机制导致piRNA位点和SV热点之间的联系(图4a): (1) piRNA位点和SV热点独立起源,它们的重叠是在共同的选择压力下通过趋同进化选择的(趋同假说);(2) SV热点首先出现,增加基因组区域进化成piRNA位点的机会(突变假说);(3)保守分子机制将piRNA生物发生与SV形成联系起来(保守假说)。为了测试SV热点是否独立产生,研究者追踪了物种间piRNA位点的突变机制。考虑到TEs和大片段重复(SDs)易于形成SVs[21, 22, 23],研究发现鸡piRNA位点与大量序列相比,TEs显著富集,但SDs显著稀缺。相反,人类粗线期piRNA位点相对缺乏TEs时[24],它们明显富含SDs[25]。尽管人类和小鼠的SDs有不同的分布[26],小鼠粗线期piRNA位点也显著富集SDs(图4b 右)。此外,研究者发现人类粗线期piRNA位点,但不是小鼠粗线期piRNA位点,与减数分裂双链断裂(DSB)热点显著重叠。研究还排除了TE转座活性导致piRNA位点不稳定的可能性,因为在鸡基因组中piRNA位点的新TE插入数量少,新TE整合位点和piRNA位点之间的距离相当远,同时在人类中已经报道了类似的反转录转座子的随机插入[27]。因此研究数据表明,鸡、小鼠和人类piRNA位点中的SV热点是独立形成的,由不同的突变机制导致(图4c)。因此,趋同进化导致SV热点和粗线期piRNA位点在鸟类和哺乳动物的基因组中同时出现。

图4 融合进化驱动SV热点和粗线期piRNA位点之间的关联
(图源:Sun YH, et al., Nat Commun. 2023)
沉默活性TEs是粗线期piRNAs的一项保守功能。为了确定是哪一种选择压力驱动趋同进化,研究重新审视了被广泛接受的观点,即哺乳动物粗线期piRNAs不参与TE沉默。研究者推断,较低的比例可能是由于SV热点的高重组率,而不是缺乏TE沉默的功能。与基因组的其余部分相比,小鼠和人类中的活性TEs(年轻且活跃的转座子都属于反转录转座子)没有从粗线期piRNA位点中减少,来自piRNA 位点的活性TEs小鼠和人类中的比例分别为1.6%和1.0%(图5a)。因此,研究测试了哺乳动物粗线期piRNA位点编码的小部分TE-piRNAs是否是TE沉默所必需的。为了避免影响前粗线期piRNAs和避免PIWI基因敲除的潜在非piRNA依赖性效应,研究者用Neurog3-cre在精母细胞中有条件地敲除了(CKO)老鼠的调控piRNA产生的基因Mov10l1[28, 29],其特异性地敲除了粗线期piRNAs,而不影响早期表达的piRNAs[30]。研究证实了先前的发现,即该突变体正常进行减数分裂并阻滞于圆形精细胞期,Mov10l1突变体的球形精子内有8±2个γH2AX病灶,而野生型球形精子细胞无任何病灶(图5b)。之前的研究将在球形精子细胞期检测到的γH2AX病灶增加归因于TE以外因素造成的DNA损伤,因为qPCR没有检测到TE表达的显著增加[41]。此研究使用RNA-seq技术发现尽管83%的TE家族(1020/1223)的表达没有改变,但是Mov10l1CKO小鼠睾丸中大多数活性TE家族(21/24;88%)显著增加(图5c) [31-35]。这个发现之前被忽略是由于以前研究的qPCR分析中使用来自TE共有序列的qPCR引物不能区分活性TE和非活性TE。研究对Mov10l1CKO小鼠睾丸中piRNAs,通过乒乓信号测量分析显示靶向这些TE的piRNA引导的对TE的降解显著减少(图5d),表明TE在Mov10l1CKO睾丸的激活是由于靶向TEs的piRNAs的丢失。这表明少量粗线期piRNAs为在粗线阶段起到了沉默活性TEs所必须。
为了测试沉默活性TEs是否也是鸡粗线期piRNAs的功能,研究者开发了一个生物信息学流程框架来定义新的TE转座插入位点。通过RNA-seq检测,这些TE家族被大量转录(图5e),并在睾丸中翻译。虽然只有2.4%的鸡粗线期piRNA位点识别活性TE,但所有活性TE家族都被粗线期piRNA降解(图5e)。因此,沉默活性TEs是粗线期piRNAs的保守功能。由于活性TEs是广泛存在、在基因组中是年轻的[36,37]、不加控制是非常有害的[38],因此抑制活性TEs的正选择作为第二种保守力量,结合高水平的局部SV突变提供进化“底物”,驱动粗线期piRNAs在羊膜动物中的快速进化,以对抗逆转录转座子的入侵和变异。
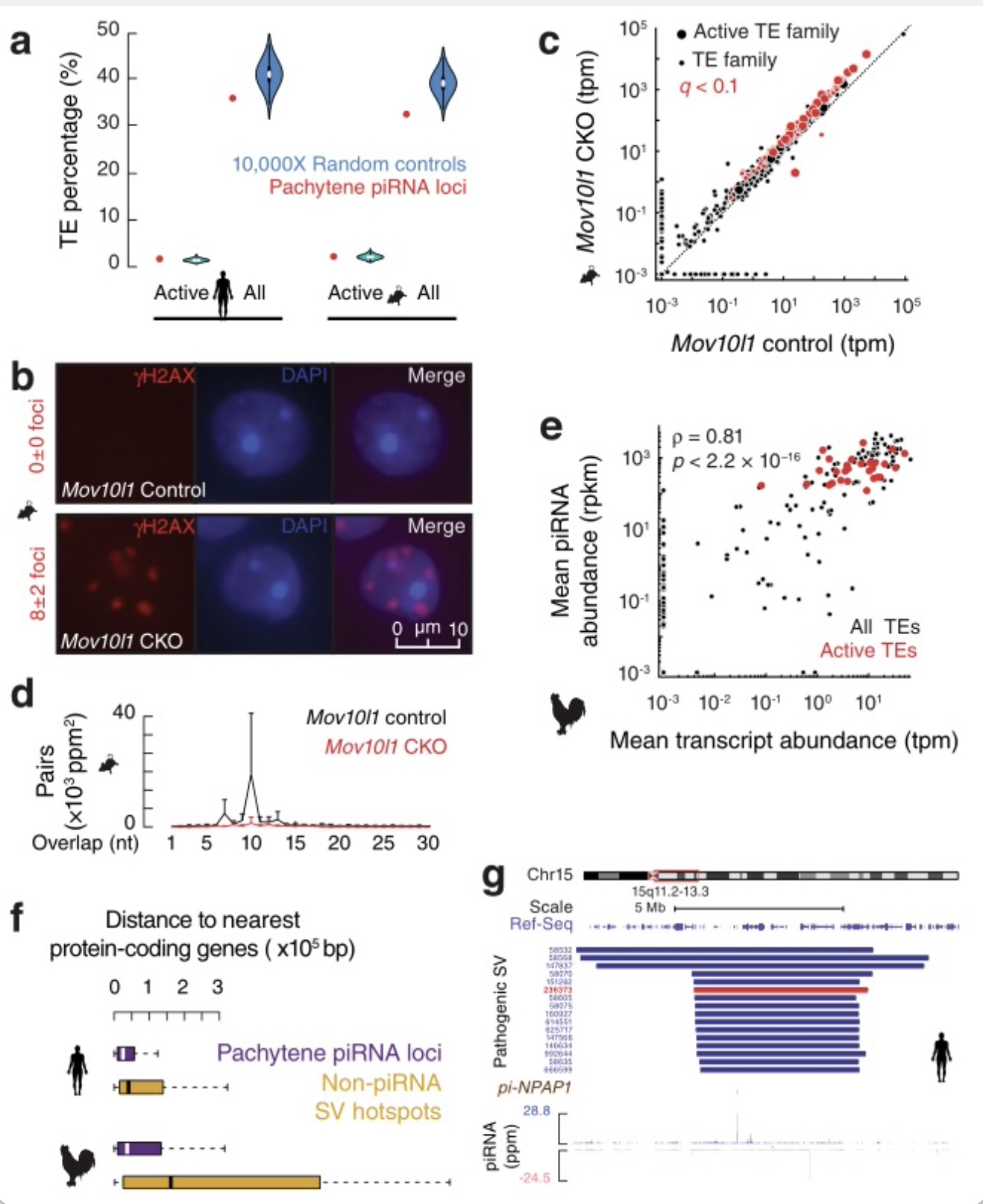
图5 沉默活性TE是驱动粗线期piRNA进化的保守功能
(图源:Sun YH, et al., Nat Commun. 2023)
提供piRNA多样性和有害SVs之间的平衡。在人类中,SVs与许多遗传性疾病有关,如发育迟缓、精神分裂症和自闭症。在人类和鸡中,SVs大多不存在于蛋白质编码区,表明它们对蛋白质功能的有害影响。研究发现在人类和鸡中,蛋白质编码基因定位在piRNA位点附近,就像它们随机分布在基因组中一样。与其他不产生piRNAs的SV热点相比,研究发现piRNA位点明显更接近人类和鸡的蛋白质编码基因(图5f),说明来源于piRNA位点的SVs比其他SV热点更容易损害蛋白质功能。研究发现那些起源于生殖细胞的致命SVs,因为它们通常破坏性太大而无法遗传给后代,与人类粗线期piRNA位点的重叠比预期的要多得多(图3e, 左)。在人染色体15q上的有害SVs,包括他和自闭症的相关性,为这种重叠提供了一个例证(图5g)。研究结果表明粗线期piRNA位点比其他不产生piRNA的SV热点更有害。因此,SV热点通过产生新的piRNAs来保护生殖细胞基因组完整性的有益功能,使得piRNA位点的SV在体细胞中的致病作用变得可忍受的。
为了测试沉默活性TEs的选择是否在进化过程中保持有害的粗线期piRNA位点,研究分析了17789个SVs,这些SVs是通过将大型猿基因组与人类基因组进行比较而确定的[55]。虽然在人类群体中这些基因组区域不再产生SVs,但这些区域在人类和类人猿的共同祖先中产生了SVs。研究发现这些历史SV区域并不富含粗线期piRNA位点(图3e)。研究比较了历史SV区域和当前人类SV热点区域的活性TE比例,发现历史SV(猿中缺失和倒位)完全没有活性TE,中位数为0%,而当前人类SV热点区域的活性TE中位数为1.5%。因此,研究结果表明,在人类进化过程中,不再能够产生piRNAs来沉默活性TEs的SV热点已经被自然选择清除了。在鸡中活跃的30个TE家族都是在鸡鸭分化后侵入鸡的基因组的,负选择解释了为什么鸡鸭共同祖先中的piRNA位点在鸡中不再存在,否则鸡基因组会积累针对古代已经失去活力的TE的piRNA位点。因此,清除有害SV热点的负选择作为piRNA位点快速更新的第三个保守驱动力。
研究发现了一个更高的局部突变率,这是羊膜动物piRNA位点快速适应的一个共同机制。虽然已知piRNA序列经历正选择以对抗TE序列[46-49],比中性突变更快固定稀有有益突变的正选择,并不是哺乳动物piRNAs快速进化的唯一原因。在研究之前,已经报道了piRNA位点的两种突变机制:(1)新的TE插入现有的piRNA位点;和(2)现有piRNA位点的拷贝数变异[5, 9]。研究发现从SV热点产生的piRNA整合了这两种突变机制,并解释了piRNA的快速产生、进化和丢失。piRNA位点的高局部突变率有利于piRNAs沉默TEs的功能,如以下五种情况所示。首先,更多样的piRNA库可以针对具有突变的TE。第二,产生TE片段的缺失将使TE无法重组离开piRNA位点,从而将截短的TE序列捕获在piRNA位点中,作为“非自我记忆”。第三,倒位从先前的有义转录方向产生反义TE-piRNAs。第四,SVs可以打破具有有害脱靶效应的piRNAs与相同前体上的必需piRNAs之间的连锁,从而允许针对有害piRNAs进行分离和选择。第五,当piRNAs靶向的TEs不再活跃时,有害的piRNA位点将在进化过程中被清除。考虑到新TEs的持续入侵,如果没有这些消除机制,粗线期piRNA位点不仅会不必要地积累,因为其他沉默机制最终会赶上来沉默古老TEs,而且还会增加有害的脱靶效应。
研究提供了一个趋同进化的例子,在两个不相关和看似冲突的过程之间产生了一种新的组合,即遗传不稳定性和保护基因组完整性的防御系统。虽然piRNAs提供了针对TEs的主要防御,但是之前的研究不知道它们如何跟上TE的高速进化。与直接起作用的保守机制相比,自然选择在进化的时间尺度上是逐渐起作用。研究推翻了保守机制的假说,即认为所有的piRNA序列都是来自共同祖先的piRNA位点的后代/旁系同源物,而是表明具有TE防御功能的piRNA位点是独立进化产生的。研究也更进一步了解粗线期piRNAs中非TE部分的功能。因为研究已经有效地排除了粗线期piRNA位点进行中性进化的可能性,这为羊膜动物中普遍存在和大量存在的非TE piRNAs提出了三种可能的解释。首先,非TE piRNA的产生对于靶向活性TE的piRNA的生物发生或功能是必需的。第二,非粗线期piRNAs具有它们自己的序列特异性功能。第三,消除这些无功能的piRNAs太昂贵或太困难。需要进一步的群体遗传学和分子生物学研究来更好地区分这些可能性。
最后,通过对人类和非哺乳动物羊膜动物的比较研究,表明SV热点保护生殖细胞基因组完整性的功能抵消了SV在体细胞中的有害作用。这支持了生殖细胞-体细胞冲突理论,该理论认为任何有助于生殖细胞存活的优势都将超过体细胞的有害作用而被选择下来[50]。研究揭示了驱动SV在基因组分布的机制。虽然以前对SVs的研究主要集中在确定它们对蛋白质编码基因的影响[51]。本研究发现确定了近百个SV热点的适应功能,主张基因组进化的研究应该从以基因为中心到以TE为中心的观点转变。
