
Complex proteins Associated with Set1 (COMPASS) 或COMPASS-like是负责催化H3K4甲基化(H3K4me)的一类复合体,并保守的存在于多个物种中,例如酵母,果蝇,哺乳动物及植物[1]。COMPASS复合体最早在酵母中被鉴定[2],仅存在一种Set1-COMPASS复合体负责催化H3K4me[1-3]。在哺乳动物中,由于H3K4甲基转移酶催化活性及辅因子的不同,COMPASS和COMPASS-like复合体被划分成了三类,分别具有不同的功能,并且多种招募COMPASS复合体到靶位点的机制也已被报道[3,4]。模式植物拟南芥中,COMPASS-like复合体的各个组分已成功鉴定[5],但关于COMPASS-like复合体的底物特异性识别机制尚未完全清楚。该研究揭示了模式植物拟南芥ATX1/2-COMPASS-like复合体位点特异性识别的新机制及JMJ28在植物免疫调控中的作用。

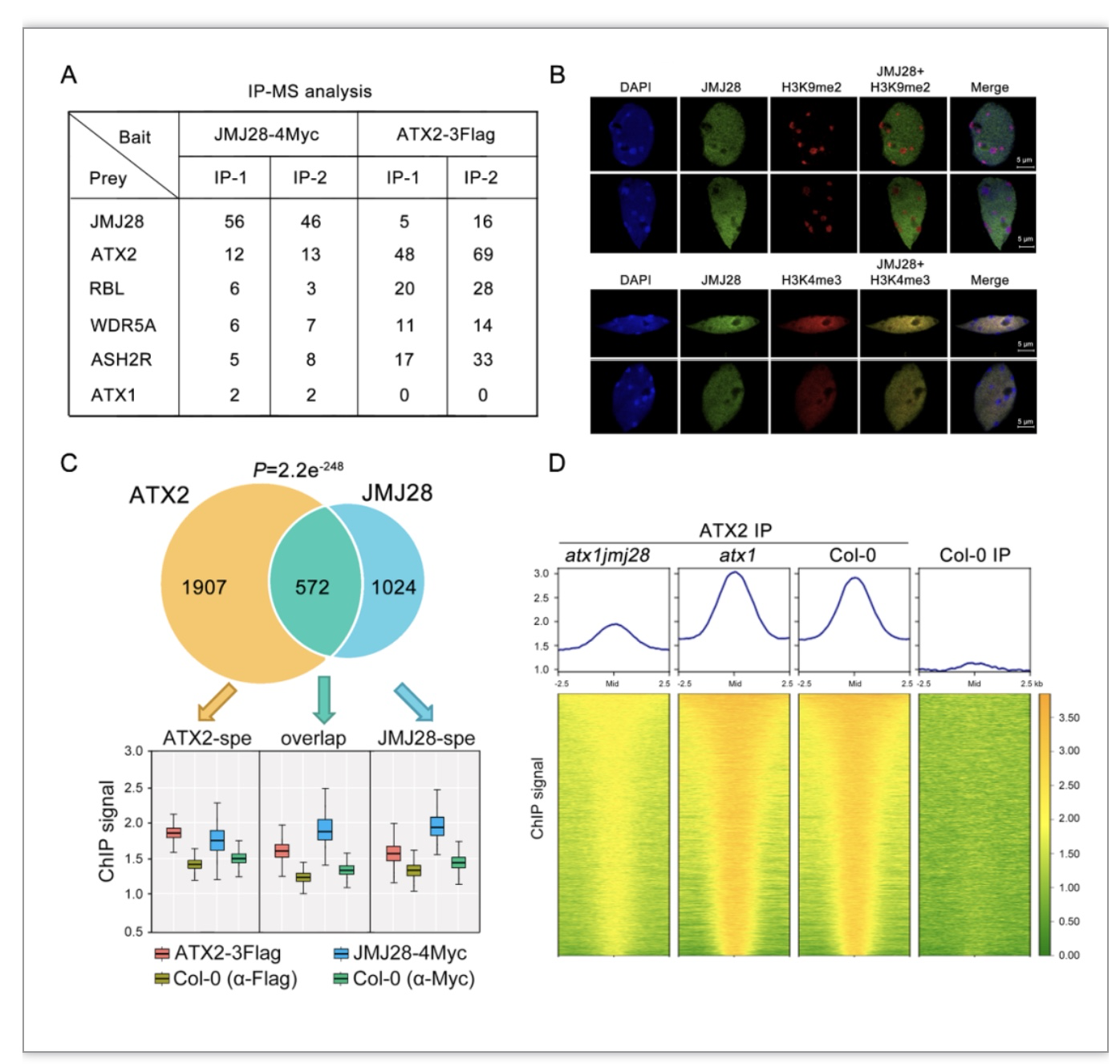
图1 JMJ28是ATX1/2-COMPASS-like复合体的互作因子并负责招募该复合体至靶位点
(图源:Xie, S. S. et al., Cell reports, 2023)
为了进一步探究JMJ28招募COMPASS-like复合体的分子机制,作者构建了JMJ28各个功能结构域的点突回补遗传学材料。研究发现,只有WRC结构域突变会影响JMJ28对靶位点的结合能力(图2A)。作者通过生信分析与EMSA实验发现JMJ28的WRC结构域负责特异的识别“YTGCAG”DNA基序(图2B-D)。当JMJ28突变后,研究发现部分位点上H3K4me3修饰水平大幅降低,基因表达也呈现显著下调。而在部分位点上,H3K4me3修饰与基因表达水平均无明显变化(图2E)。作者通过进一步探索,发现ATX1/2-COMPASS-like复合体在部分位点上的调控可能与其他类型COMPASS-like复合体功能冗余(图2F)。有趣的是,作者还发现JMJ28参与负调节植物免疫(图2G)。JMJ28自身的基因表达在响应病原体侵染时显著下调。此外,其介导的H3K4me3的沉积也参与了多种免疫基因的转录调控。综上所述,该研究揭示了植物中COMPASS-like复合体实现底物特异性的一种分子机制(图3)。
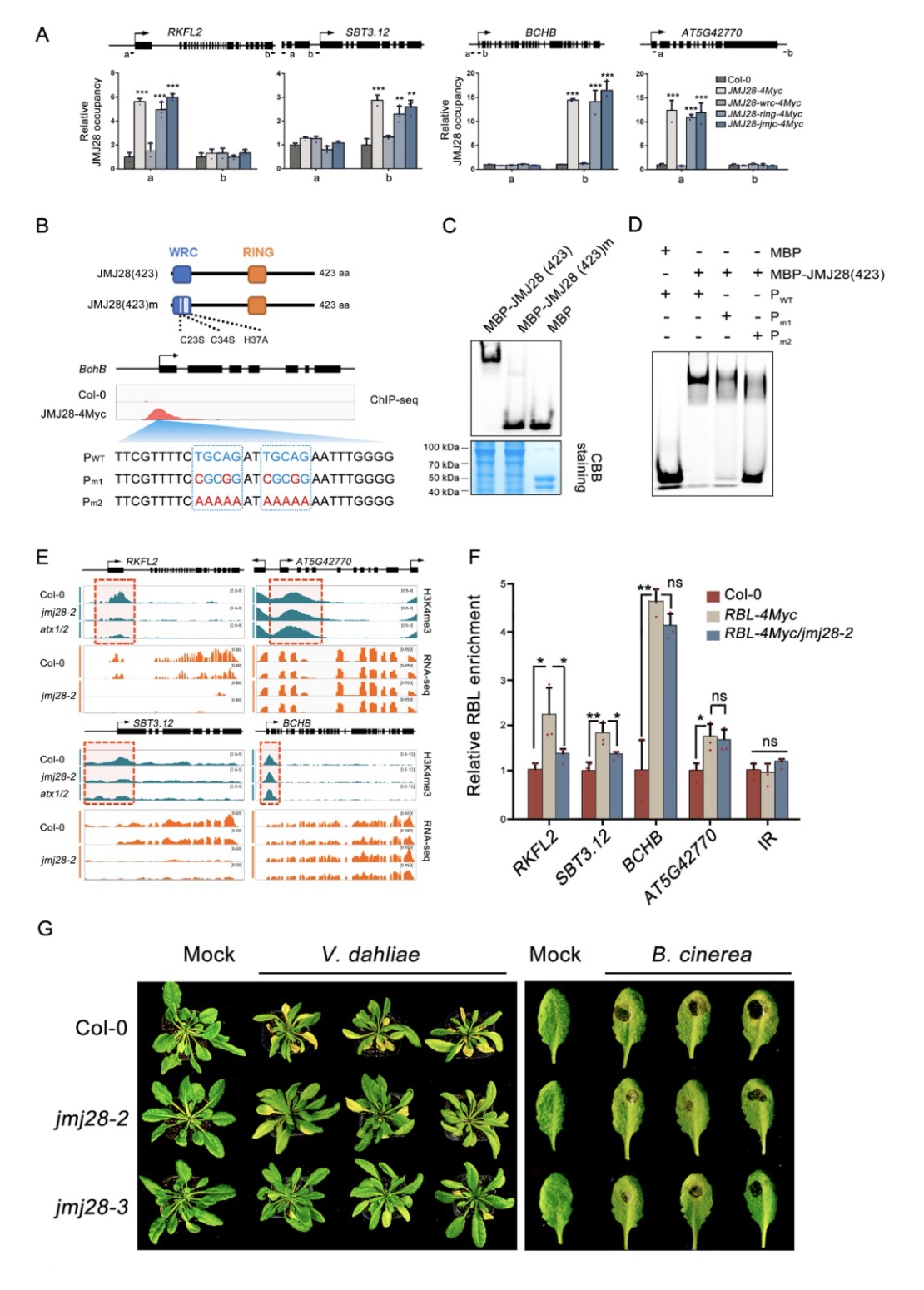
图2 JMJ28介导靶位点上H3K4me3修饰的沉积并参与植物免疫调控
(图源:Xie, S. S. et al., Cell reports, 2023)
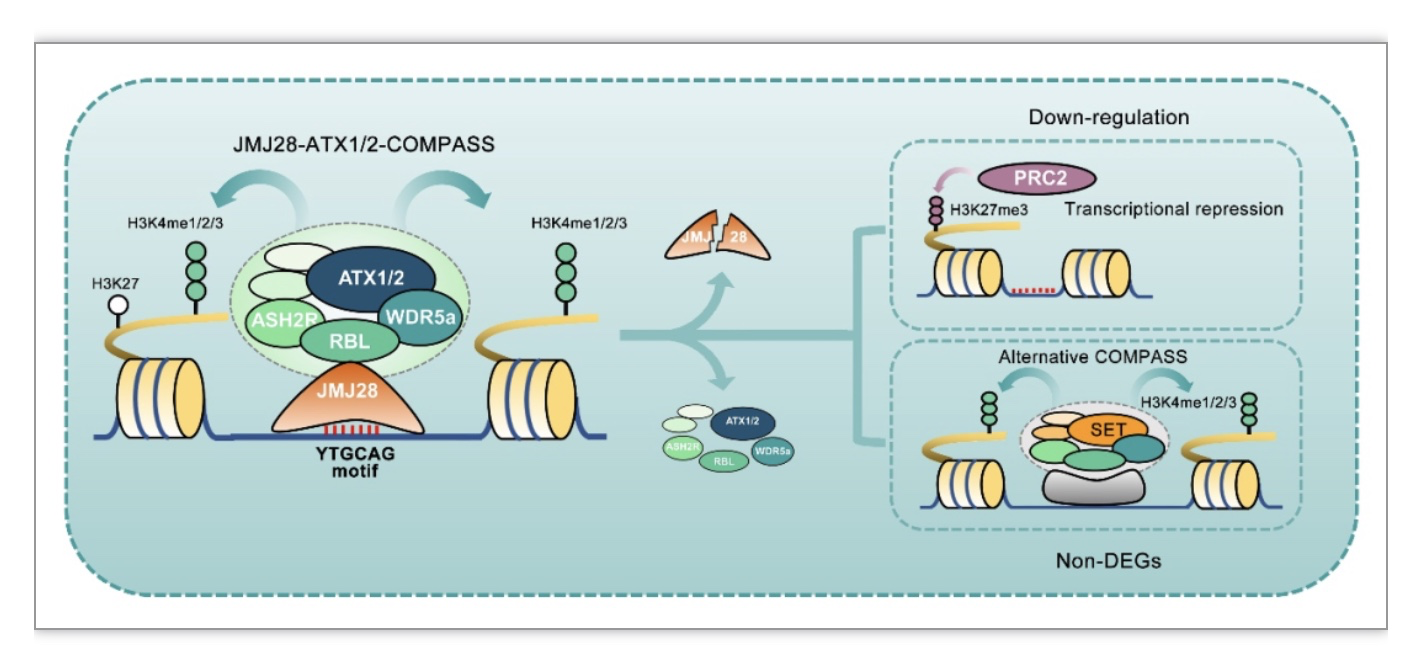
图3 JMJ28介导的ATX1/2-COMPASS-like复合体底物特异性识别的分子模型
(图源:Xie, S. S. et al., Cell reports, 2023)
由于COMPASS-like复合体之间存在广泛的功能冗余,以及与其他甲基转移酶相比,ATX1/2对H3K4me3的贡献相对较小,导致鉴定出的受JMJ28特定靶向的位点数量并不多。另外,目前尚不清楚JMJ28所识别的“YTGCAG”DNA基序是否也被其他类型COMPASS-like复合体的相互作用因子识别,甚至被其他生物使用。这些机制都有待未来进一步探究。
