18101298214
公司首页
实验手册
小分子化合物
细胞培养和检测
定制服务
文献资料
联系我们
订购指南
公司首页
实验手册
小分子化合物
细胞培养和检测
定制服务
文献资料
联系我们
订购指南
首页
实验手册
发现组蛋白变体H1.2调控脂肪组织产热
发布日期:2023/9/9 12:13:00
脂肪细胞
分为白色脂肪细胞和棕色脂肪细胞,但是在寒冷暴露或β3-肾上腺素刺激下,第三种形式的脂肪细胞—米色脂肪细胞(具有棕色脂肪细胞样产热表型)在白色脂肪组织(WAT)中被诱导,这一过程被称为棕色化。诱导具有产热能力的棕色和米色脂肪细胞的数量可激活能量消耗,防止高脂饮食(HFD)诱导的肥胖,改善高脂血症,并改善小鼠的全身葡萄糖稳态和胰岛素敏感性。
因此,了解调控棕色化的机制将有助于开发针对肥胖和相关代谢并发症的新疗法。
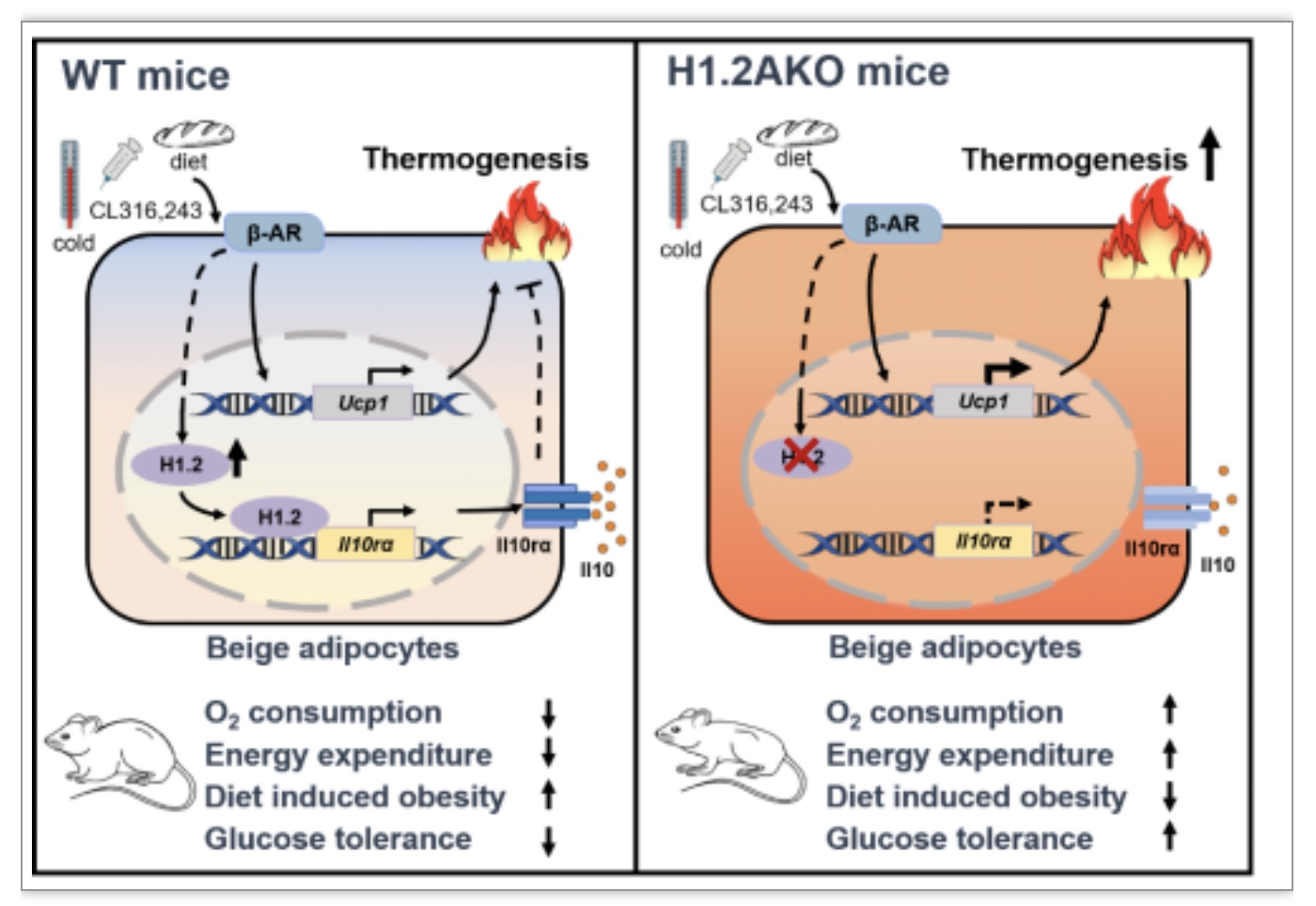
该研究发现连接组蛋白变体H1.2在产热脂肪中高度表达,并且是米色脂肪细胞激活的刹车分子,能够感应产热刺激并通过调控Il10rα表达来抑制白色脂肪组织棕色化,H1.2-Il10rα信号轴在脂肪组织产热中的调控作用对研究肥胖及其相关代谢疾病的发病机理有重要的参考价值。
研究材料
在这项研究中,研究人员利用H1.2
flox/flox
小鼠与adiponectin-Cre小鼠杂交得到了C57BL/6背景的脂肪细胞特异性H1.2敲除小鼠(H1.2AKO),其中使用的H1.2
f
lox/flox
小鼠是由赛业生物提供的。此外研究者还运用了原代脂肪细胞、人皮下脂肪样本等进行研究。
研究方法
作者采用了多种实验技术,包括各种组织学染色、WB、qPCR、转录组学测序(RNA-seq)、染色质免疫共沉淀(ChIP)、染色质免疫共沉淀测序(ChIP-seq)和双荧光素酶报告基因实验等。
技术路线
01确认了组蛋白H1.2在脂肪组织中的表达量响应产热刺激
02通过H1.2AKO小鼠和H1.2过表达小鼠确认H1.2是产热的负调节因子
03探究H1.2调控能量代谢的机制
04通过HFD喂养模型证明H1.2是肥胖的重要靶点
05通过脂肪组织原位注射回补Il10rα证明H1.2对代谢的改善作用依赖于Il10rα
研究结果
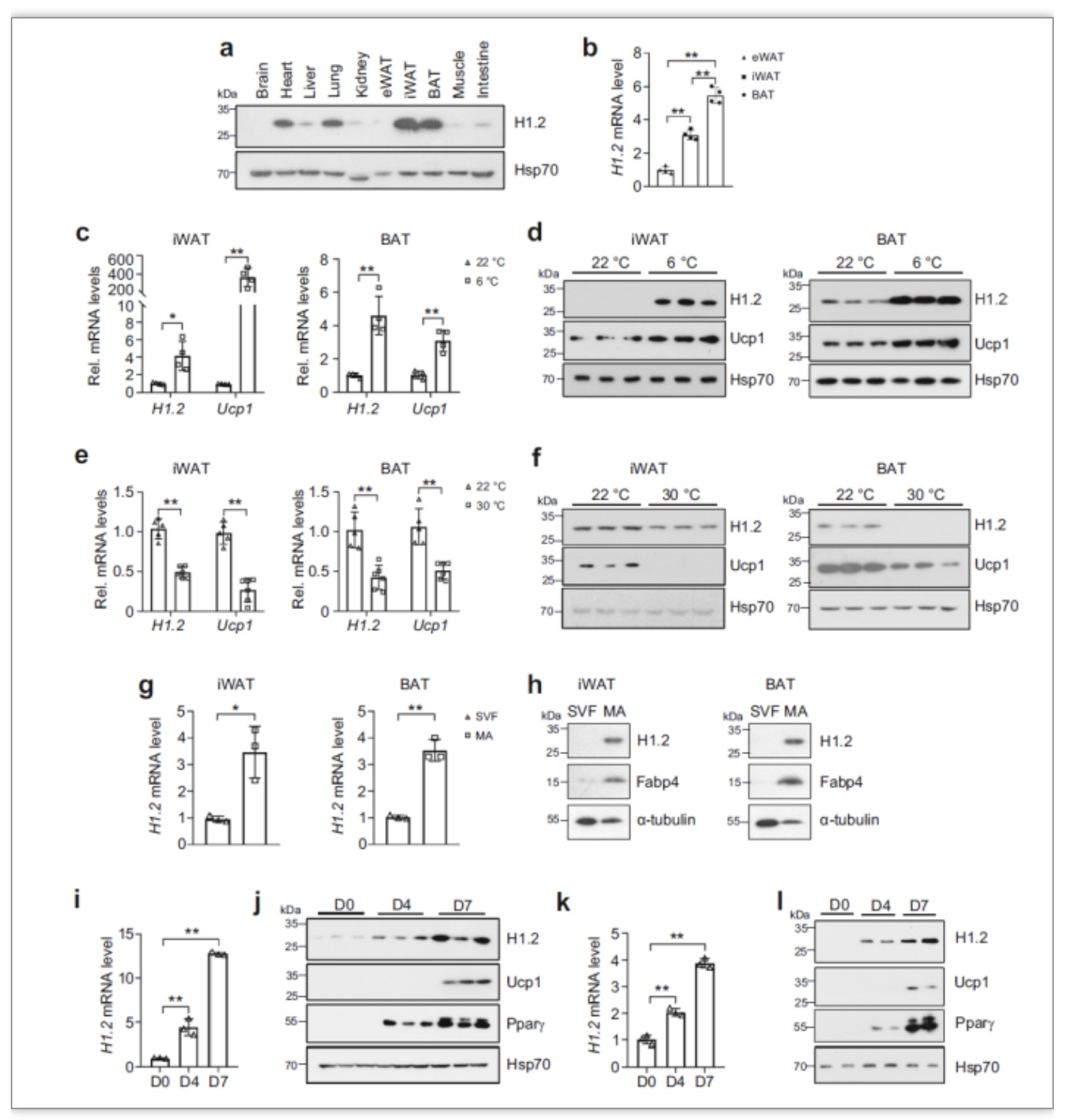
图1 H1.2在米色和棕色脂肪细胞中高表达并响应温度刺激[1]
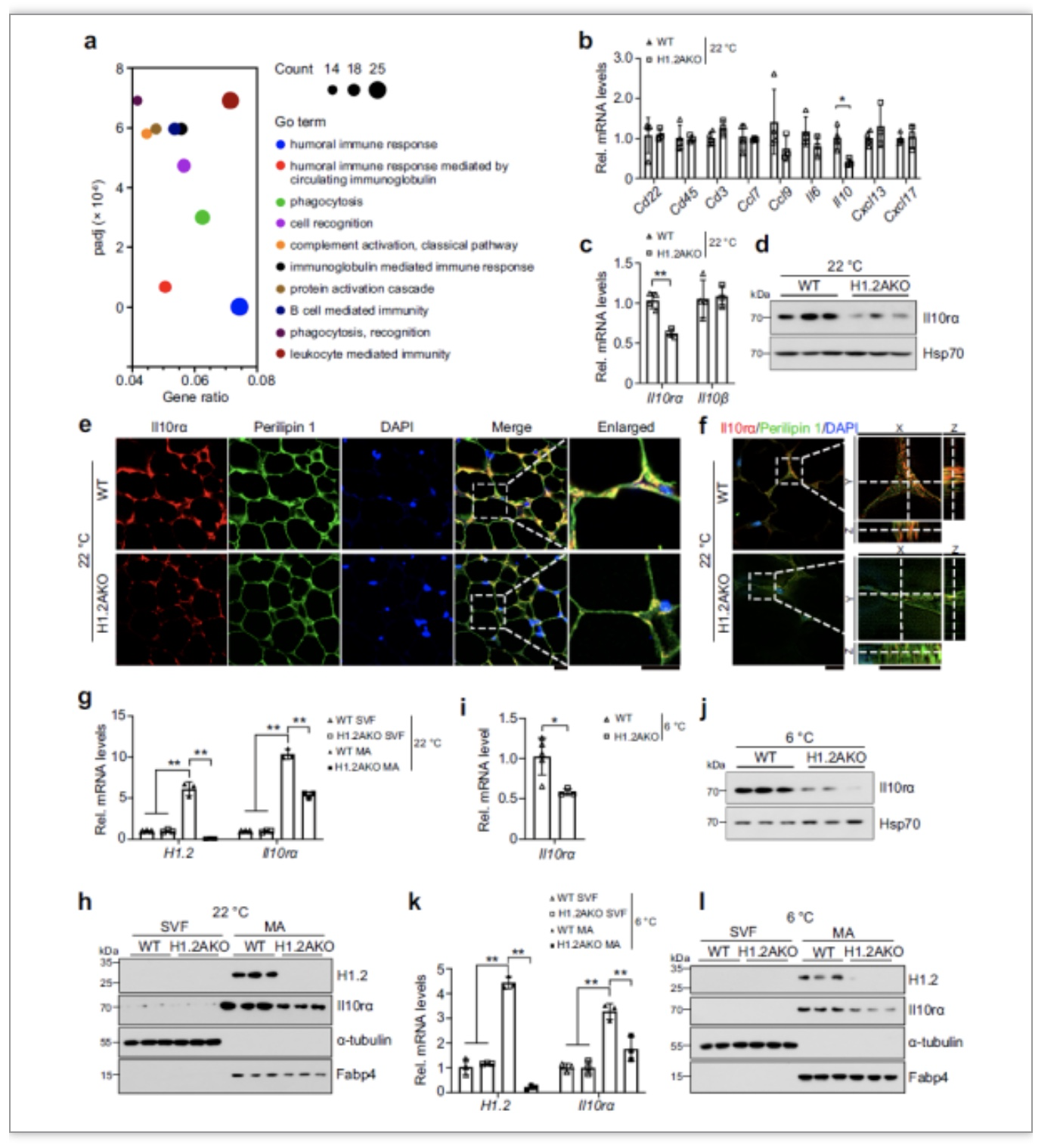
首先,作者检测了12周龄的C57BL/6雄鼠的心、肝、肾、肺、脑、脂肪、肌肉、小肠等10个组织中H1.2的表达情况。结果表明,在各个组织中H1.2的表达差异较大,在腹股沟处白色脂肪(iWAT)和肩胛骨处棕色脂肪组织(BAT)中表达最高,并且转录水平与蛋白表达水平相似。接下来,为了探究H1.2的表达量是否和产热状态有关,作者分别在冷刺激和热中性温度下检测小鼠iWAT和BAT中H1.2的表达量,
发现冷刺激下H1.2表达量上升,热中性温度下,H1.2表达量下降。
之后作者分离了iWAT和BAT中的血管基质层细胞(SVF)和成熟脂肪细胞(MA),发现H1.2主要在成熟脂肪细胞中表达。检测体外分化的米色脂肪细胞和棕色脂肪细胞中H1.2的表达量,也发现,
H1.2在成熟的脂肪细胞中表达量更高,如图1所示。
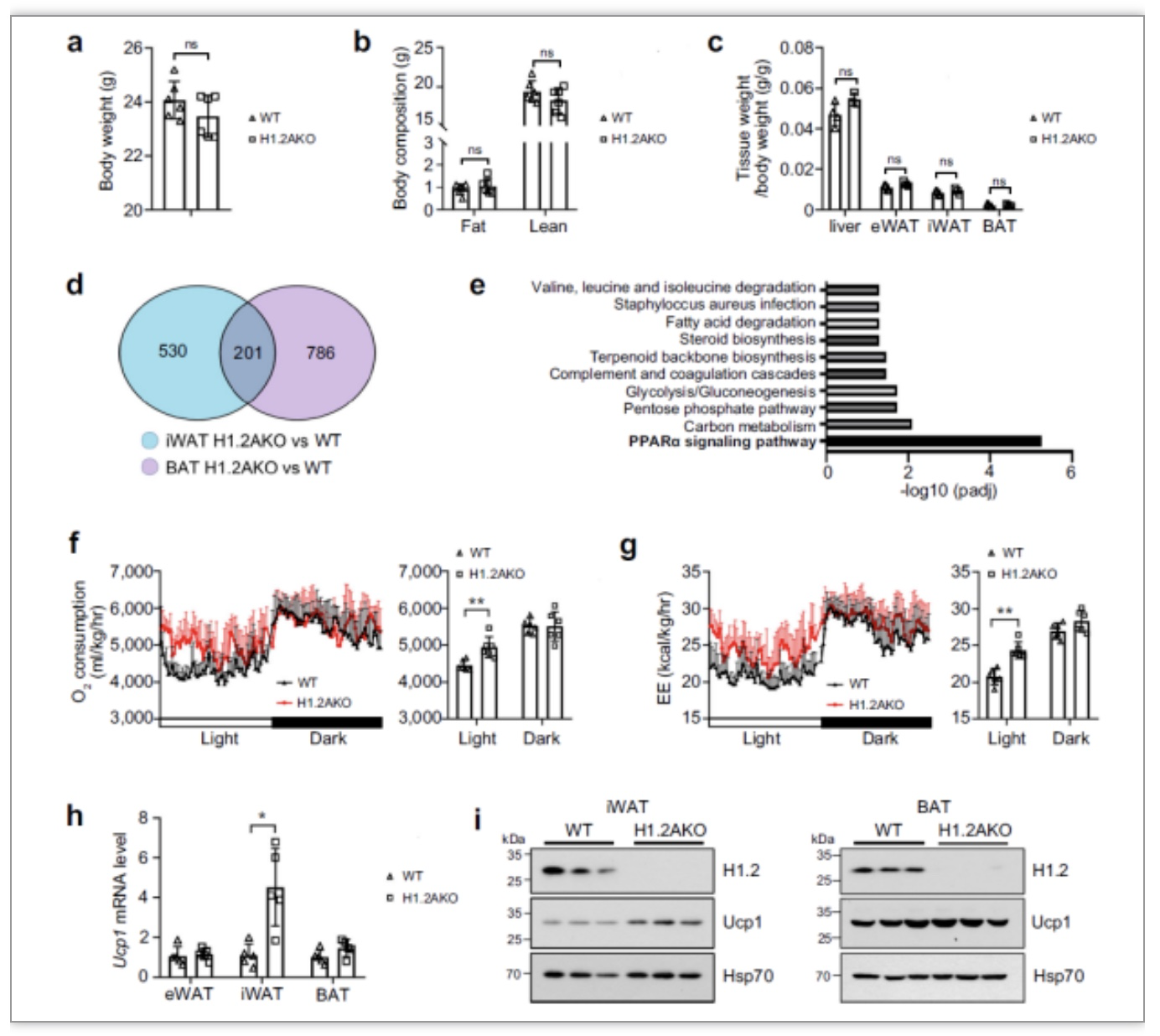
图2 H1.2AKO小鼠能量消耗更高[1]
为探究H1.2的生理功能,利用H1.2AKO小鼠进行了RNA-seq,发现变化基因集中在PPARα信号通路(其中,H1.2
flox/flox
小鼠由赛业生物提供)。代谢笼监测小鼠的耗氧量,发现在白天,H1.2AKO小鼠的耗氧量与WT小鼠相比显著性升高,说明H1.2AKO小鼠具有较高的基础代谢率。此外,
在H1.2AKO小鼠iWAT中产热相关重要基因Ucp1的转录水平和蛋白水平都有显著性升高,而在BAT中没有明显差异,如图2所示。
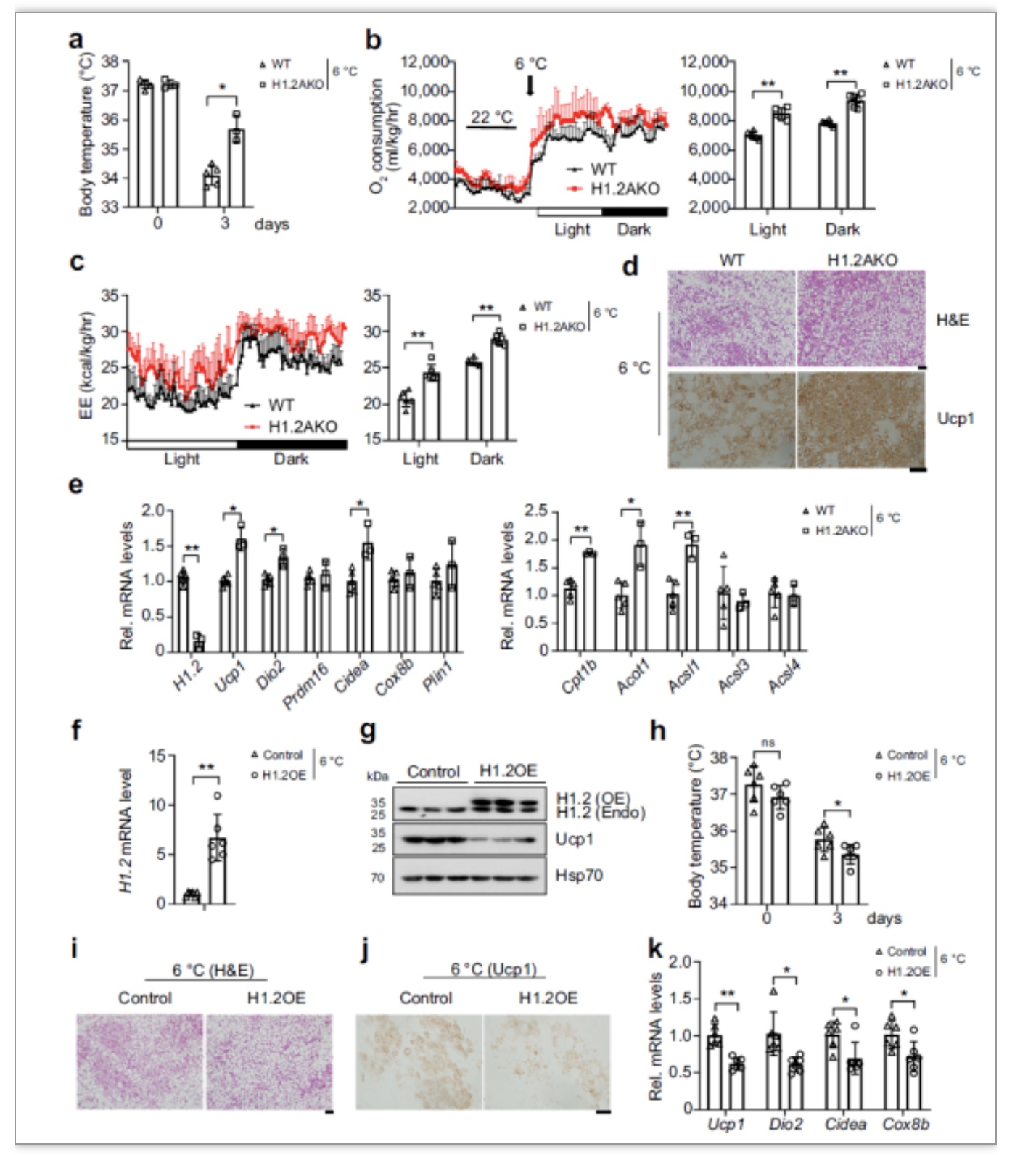
图3 H1.2在冷刺激下调节iWAT转化[1]
既然H1.2AKO小鼠的产热基因表达更高,接下来作者将小鼠放于6℃的环境中,检测H1.2AKO小鼠和WT小鼠在冷刺激下的产热情况。
发现H1.2AKO小鼠对冷刺激更耐受,而在iWAT中过表H1.2(H1.2OE)的小鼠对冷刺激更不耐受,如图3所示。
图4 Il10rα在H1.2AKO小鼠的成熟脂肪细胞中表达量下调[1]
为了寻找H1.2A调控iWAT产热的生理机制,研究人员对WT小鼠和H1.2AKO小鼠iWAT中的RNA-seq结果进行了分析。将差异基因进行GO(Gene Ontology)信号通路富集,发现变化最明显的基因富集在体液免疫方面,包括细胞识别、白介素介导的免疫应答、B细胞介导的免疫反应等,暗示H1.2可能通过调控iWAT中的免疫细胞来影响产热。
qPCR检测发现在H1.2AKO小鼠的iWAT中Il10的转录水平有显著性下调,Il10的受体Il10rα的转录水平也有显著性下降,而Il10rβ的转录水平没有差异。
为了进一步验证H1.2AKO小鼠的iWAT中Il10rα的降低发生在成熟脂肪细胞,还是其他细胞类型,作者从WT和H1.2AKO小鼠iWAT中分离出相应的血管基质层(SVF)细胞和成熟脂肪细胞(MA),分别检验其中Il10rα的表达量。
结果表明,H1.2AKO小鼠iWAT中Il10rα的下降主要发生在成熟脂肪细胞。冷刺激下也可以看到同样的现象,如图4所示。
图5 H1.2AKO小鼠抵御HFD喂食诱导的肥胖和代谢紊乱[1]
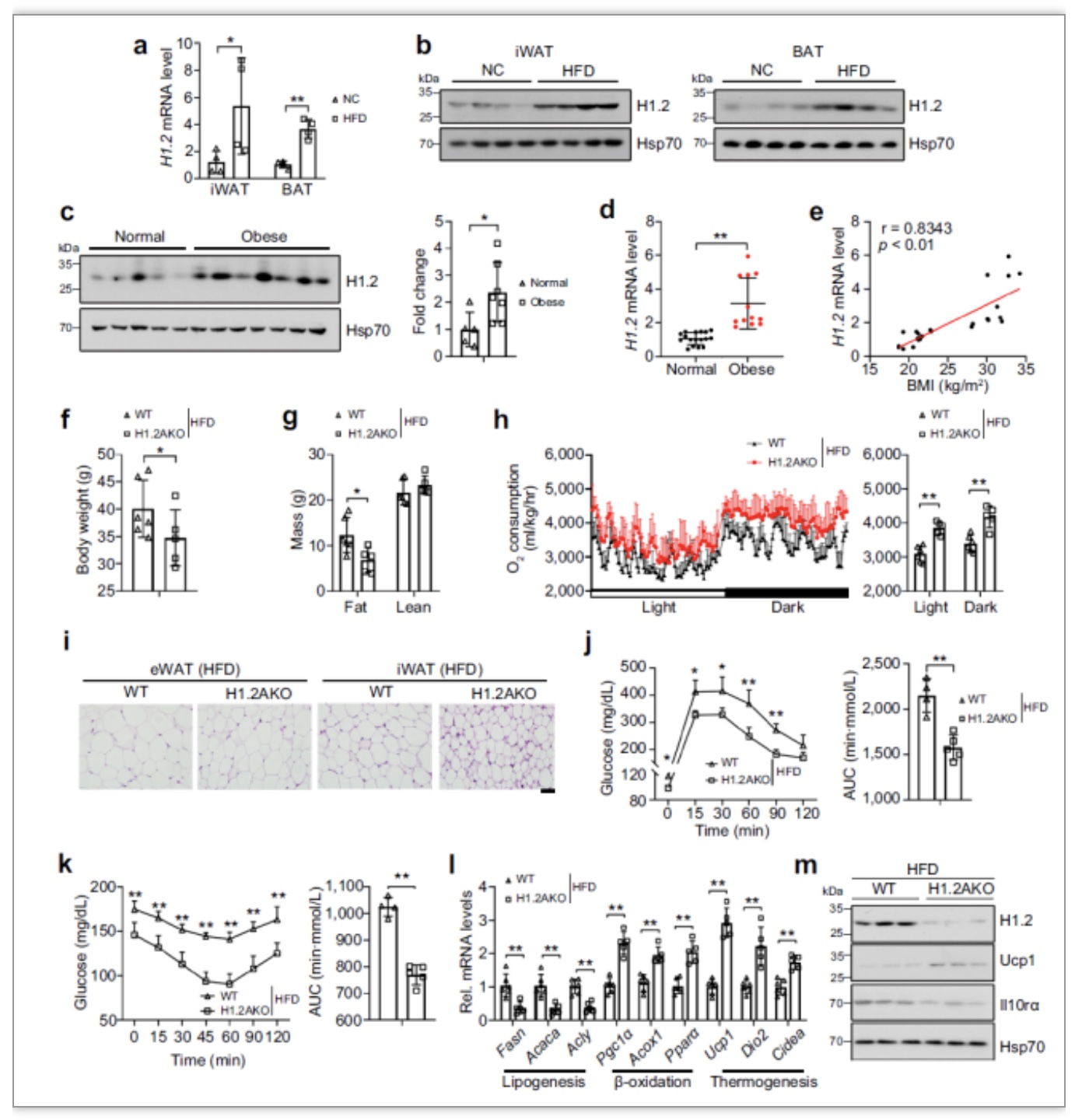
作者还发现,不论是在肥胖的小鼠还是肥胖的人群中,H1.2的表达量都显著升高。并且,
在高脂食物食物喂养下,H1.2AKO小鼠体重更低,脂肪更少,氧消耗更高,糖代谢更好,产热基因的表达水平也更高,伴随着更低的Il10rα表达量,如图5所示。
图6 H1.2AKO小鼠对代谢的改善作用依赖Il10rα[1]
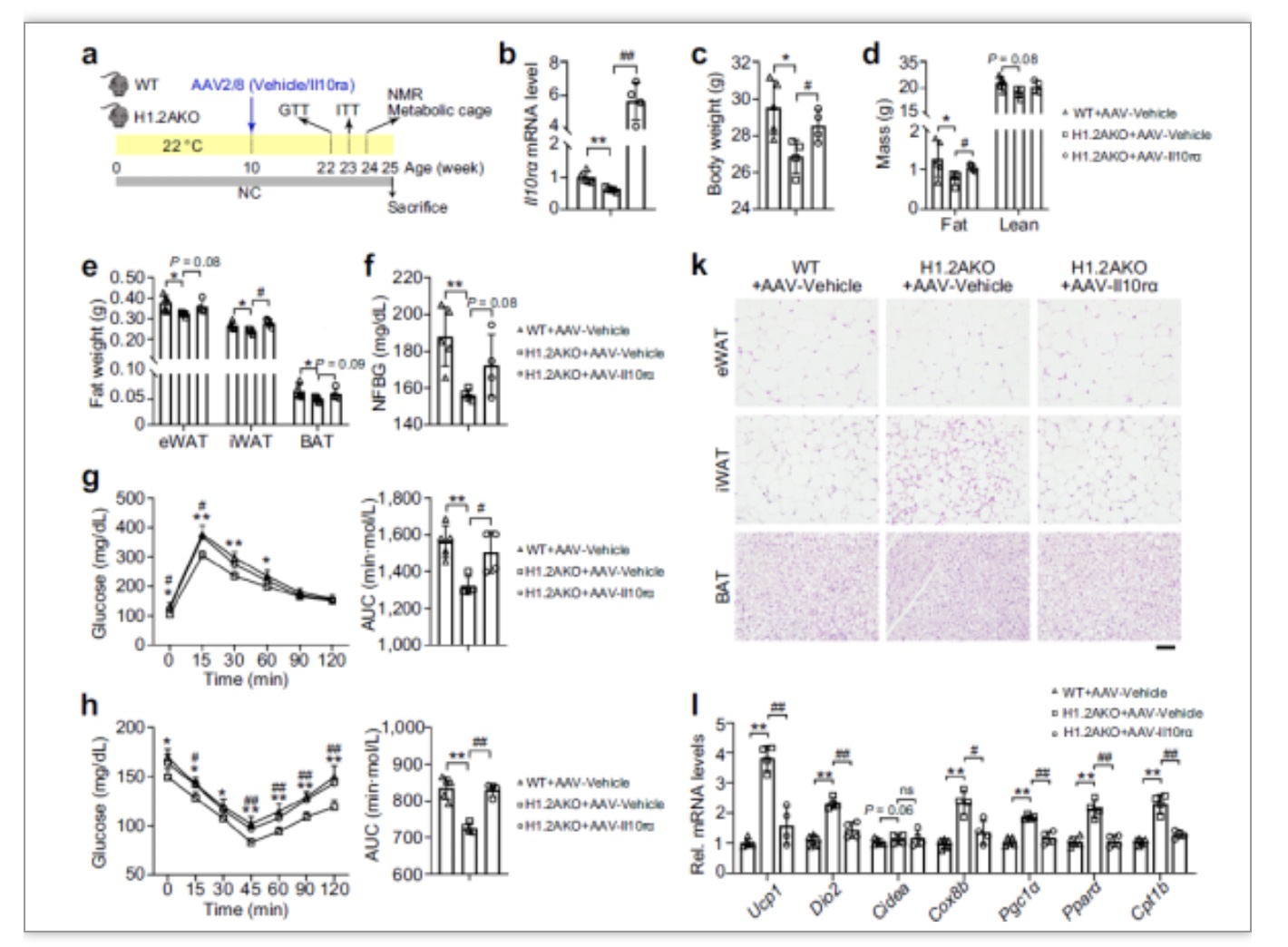
为了证明H1.2AKO小鼠对代谢改善的作用依赖于Il10rα,在iWAT中原位注射腺相关病毒AAV-Il10rα,发现过表达Il10rα后抵消了H1.2AKO小鼠脂肪积累更少,改善糖代谢的作用,
说明H1.2AKO主要是通过调控Il10rα的表达来影响产热和代谢,如图6所示。
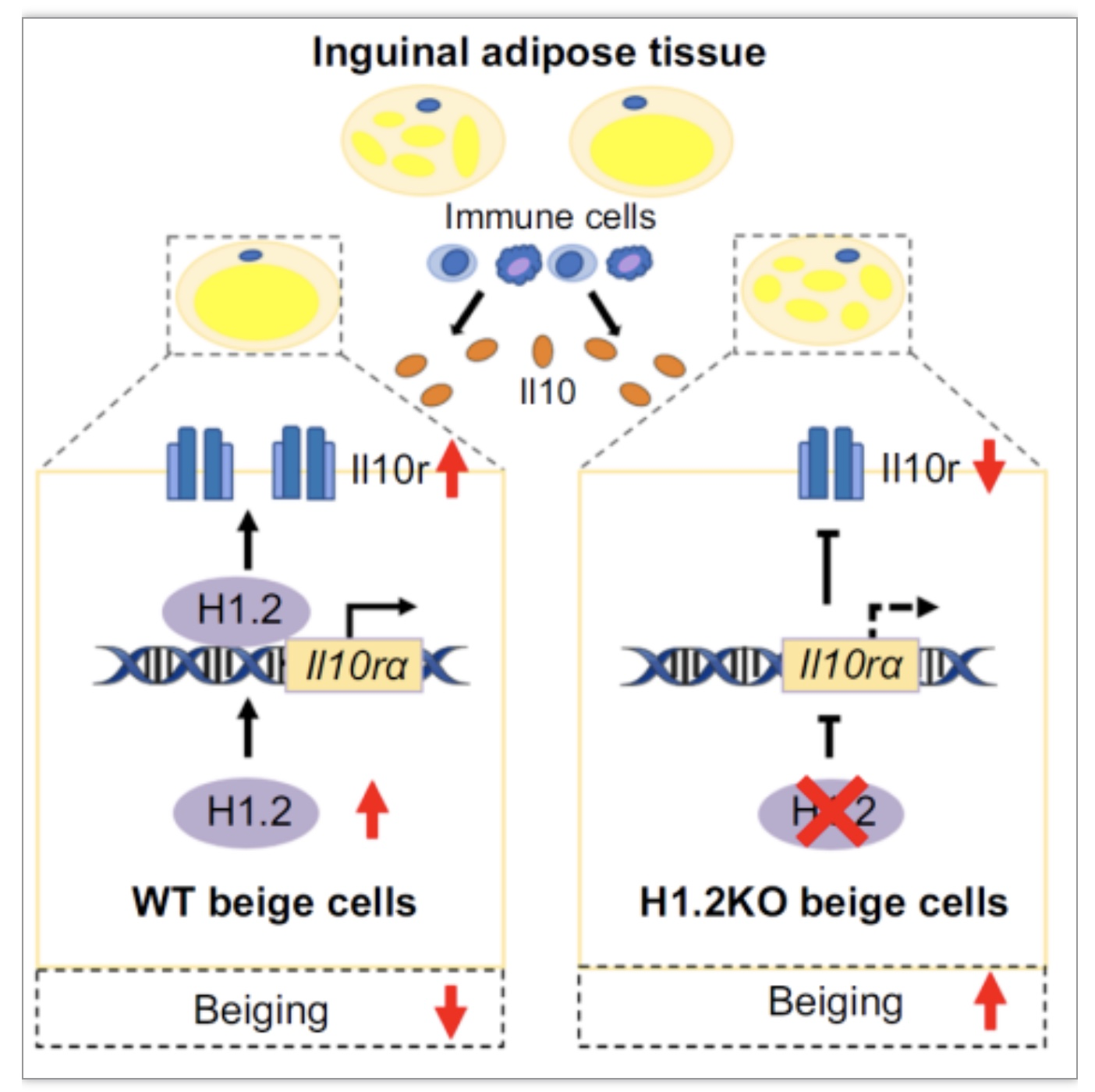
图7 WT米色脂肪细胞与H1.2缺失的米色脂肪细胞中Il10rα水平变化[1]
文章结论与讨论,启发与展望
该研究阐明了组蛋白变体H1.2在脂肪能量代谢中的功能机制,即H1.2AKO可以通过调控Il10rα的表达来影响产热和代谢。该研究中H1.2-Il10rα信号轴在脂肪组织产热中的调控作用对研究肥胖及其相关代谢疾病的发病机理有重要的参考价值。
上一篇:
通过线粒体自噬调控肾缺血/再灌注损伤
下一篇:
评述抑癌基因TP53的新进展
已经到最底了
技术支持:
库价化学
Copyright © 2024北京螽斯羽生物有限公司 备案号:
京ICP备2023018288号-1