
转录因子(TFs)与目标基因启动子/增强子中的特定DNA响应元件的结合是调节基因转录以及随后的细胞过程的关键事件。为了使基因正常表达,TFs必须在正确的时间和地点选择性地相互作用,并与特定的DNA序列和辅助因子组装成高阶复合物[1-2]。在真核生物基因组中,转录因子从基因组内大量的潜在结合位点中准确地结合一小部分相关结合位点的能力是建立转录调控的基础。之前大量结构研究提供了关于转录因子NF-kB如何识别其目的DNA (κB DNA)结合位点的宝贵信息[3]。然而,转录因子NF-kB如何识别和区分序列相似但功能不同的DNA序列还不是很清楚。
该课题组之前的研究发现p52:p52同源二聚体可以感知κB DNA序列中央位置从G/C到A/T的单碱基突变。p52:p52同源二聚体能同时结合两种κB DNAs;但只有在DNA中心是G/C的情况下,p52:p52同源二聚体才能与其特定的辅助因子Bcl3结合(形成p52:p52:Bcl3复合物),并通过招募组蛋白乙酰转移酶激活转录。当与序列中心为A/T的DNA结合时, p52:p52:Bcl3复合物将会招募组蛋白去乙酰化酶而抑制基因转录。为何单个不和蛋白直接接触的核苷酸的突变竟然会对转录的选择性产生如此巨大的影响,其分子机制至今仍不清楚。该研究揭示了κB DNA与NF-κB复合物的新型构象,并指出了由DNA构象以及动态决定的结合动力学在调控NF-κB的转录激活方面的重要性。

在过去的25年中,已经报道了多种κB DNA与NF-κB二聚体的复合物结构。在所有这些结构中,DNA序列是以A/T为中心的κB位点。而PSel-κB DNA(5′-GGGGTGACCCC-3′)(红色为中心碱基,下划线为±1位置的碱基)是一个已知由p52:p52:Bcl3复合物特异性调节的天然结合位点,其序列不仅在中心位置而且在两个侧翼位置都与经典的κB位点不同。研究者们对中心和侧翼的碱基进行了突变,以产生PSel(中心碱基突变成A/T)(5′-GGGTAACCCC-3′)和(-1/+1位置的碱基交换)(5′-GGGAGTCCCC-3′)DNA。Vivien Wang团队发现与天然的PSel-κB DNA相比,PSel(A/T突变体)降低了转录活性(约2倍),而PSel(-1/+1碱基交换)和MHC-κB DNA转录活性都急剧降低(图1B)。这些结果表明,中心区域所有三个位点上的碱基特征在决定p52:p52:Bcl3复合物的转录活性方面至关重要。随后该团队解析了p52:p52同源二聚体与三种不同的PSel-κB DNAs复合物的晶体结构。研究者发现与以前已知的以A/T为中心的NF-κB-DNA复合物的结构相比,三个PSel-κB DNAs在围绕中心位置0(-1到0和0到+1)的两个碱基上表现出明显的小沟变宽,宽度约为7.5 Å(图1E-G1)。先前研究的κB-33(5′-GGAAATTTCC-3′)和κB-55(5′-GGGAATTCCC-3’以A/T为中心的κB DNA),与理想的B型DNA相比,它们在结合和自由状态下的小沟都有明显的挤压(图1I)。挤压小沟宽度是所有与NF-κB二聚体结合的以A/T为中心的κB DNA的一个共同特征,这与本结构中看到的PSel-κB DNA的小沟宽度明显不同。
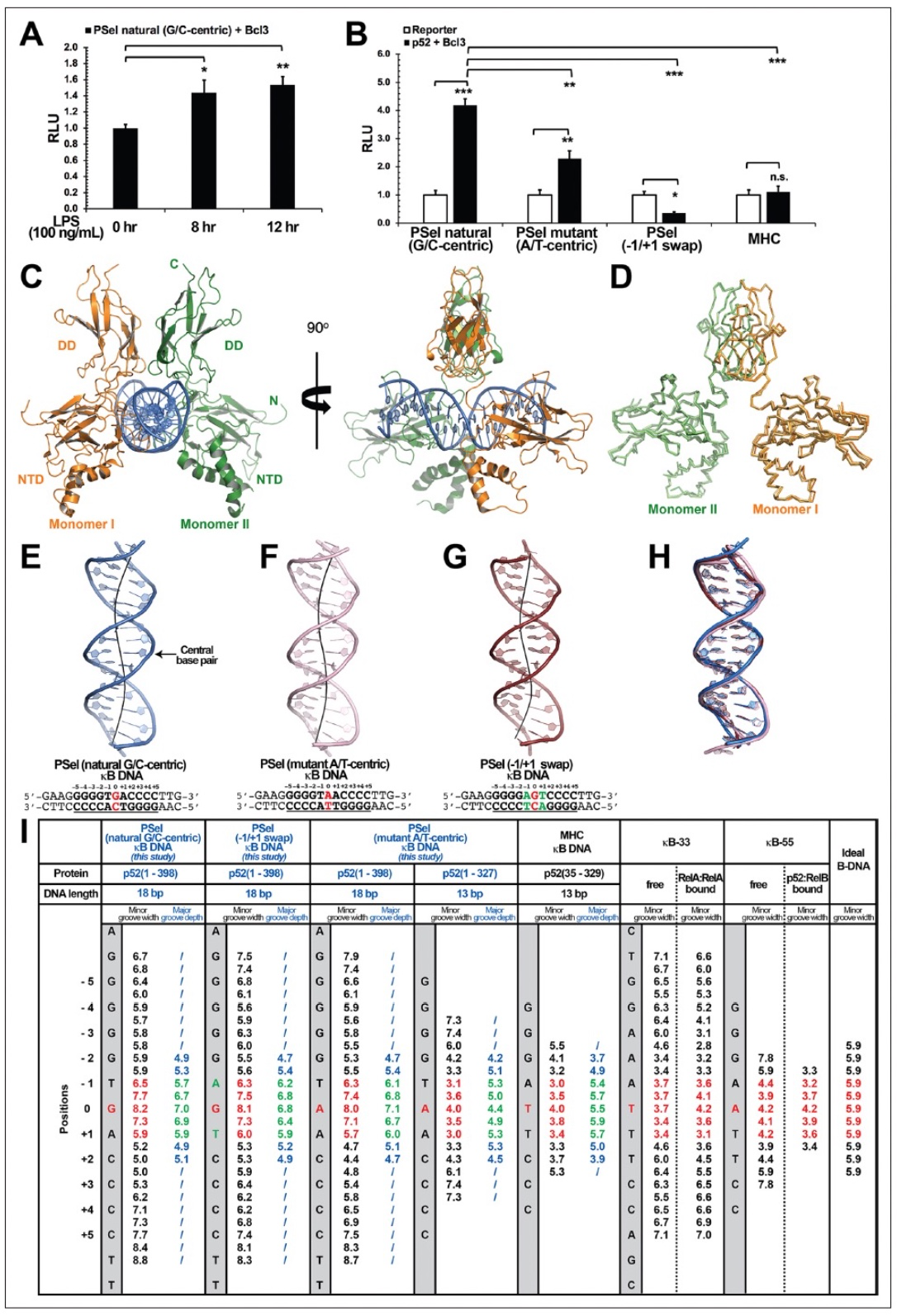
图1: PSel-κB DNA突变体与p52:p52同源二聚体的复合体晶体结构
(图源:Pan et al.,eLife,2023)
为了研究在目前的复合物中观察到的PSel-κB DNA变体的小沟宽度是由蛋白质诱导的还是DNA序列固有的,香港中文大学合作者对四个κB DNA的自由形式和与p52同源二聚体复合物形式进行了微秒级的分子动力学(MD)模拟。结果表明,其中三个PSel-κB DNAs变体都显示出较宽的小沟,而MHC-κB DNA有一个狭窄的小沟(图2)。在与p52:p52同源二聚体结合后,所有DNA的中心小沟都变窄了,和在自由形式下看到的一致(图3)。为了适应DNA中央小沟的宽度,与-1/+1交换DNA结合的p52:p52亚基采用封闭的构象,其中与中央小沟结合的两段似乎夹住了DNA,与它们和天然G/C为中心的DNA结合时的更开放构象相反;与A/T突变体DNA结合的p52亚基处于上述两类构象之间。值得注意的是,研究者发现p52:p52亚单位的NTD在与突变体A/T中心和-1/+1互换DNA结合时,可以在中央小沟中进行跨链接触(图3A)。此跨链接触,虽然在三种蛋白质-DNA复合物中都存在,但在-1/+1碱基交换和A/T突变体 DNA中出现频率较高。这种差异表明,p52:p52同源二聚体的多种动态构象令它能识别三种PSel-κB DNAs的特定小沟的几何形状。
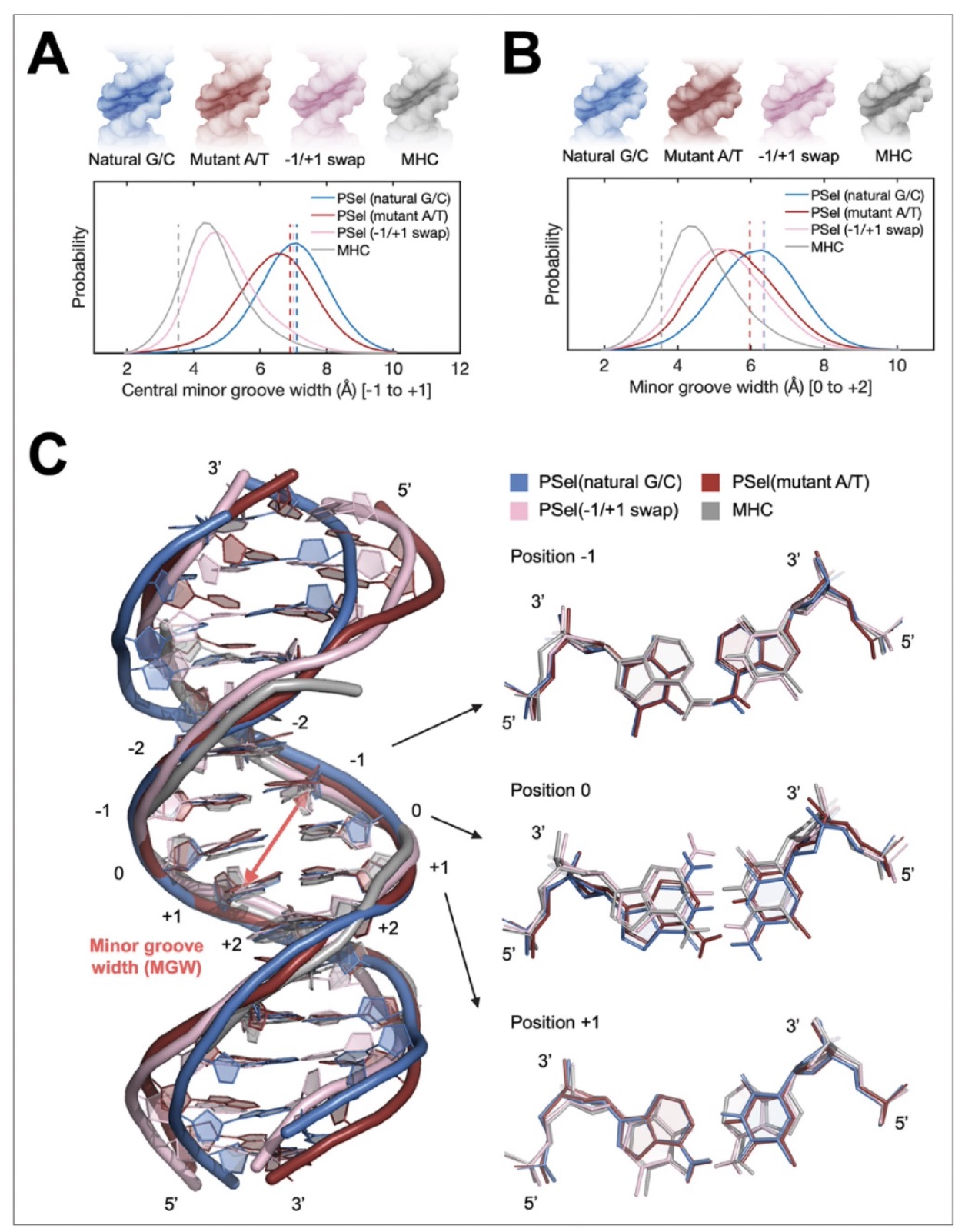
图2: 自由形式的κB DNA的分子动力学模拟
(图源:Pan et al.,eLife,2023)
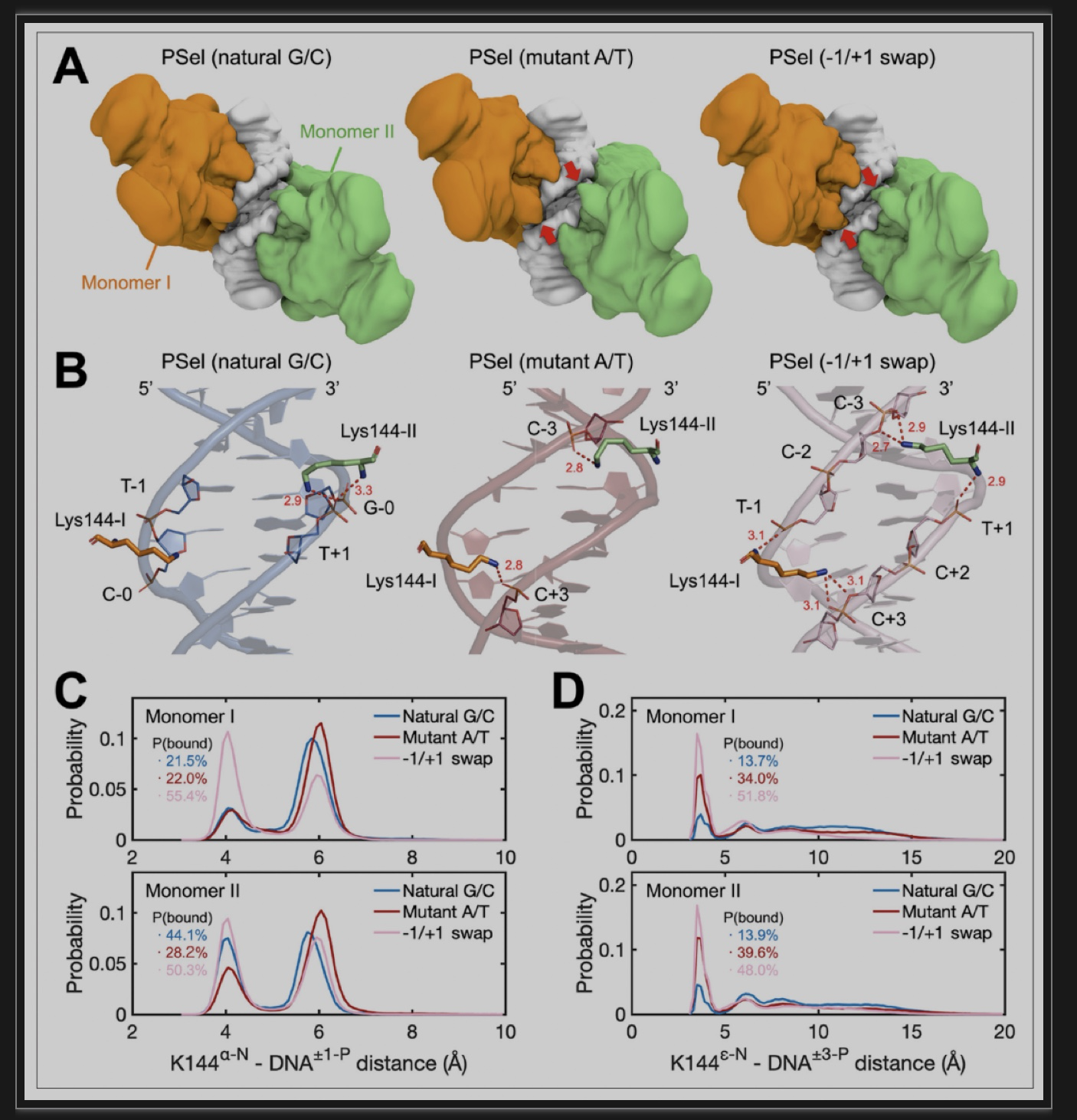
图3: (p52:p52)-DNA复合物的分子动力学模拟
(图源:Pan et al.,eLife,2023)
然而,上述结构分析没有提供(p52:p52)-DNA复合物的构象状态与转录输出之间的明显关联性。因此该团队还测试了p52:p52同源二聚体是否以不同的机制和/或亲和力与天然的G/C为中心的PSel-κB DNA、A/T为中心的突变PSel-κB DNA、-1/+1交换的PSel-κB DNA,以及MHC-κB DNA结合。结果显示,p52:p52与所有三种PSel-κB DNAs变体的结合亲和力(Kd)相似,而它与MHC-κB DNA的结合更紧密(图4)。然而,p52与天然G/C为中心的PSel-κB DNA的结合与熵(ΔS)的大量增加和焓(ΔH)的适度降低有关。另一方面,与MHC和A/T突变体的PSel DNA的结合显示出更大的焓值下降。这些结果表明,p52:p52同源二聚体与以G/C为中心的κB DNA的结合更受熵的影响,而与以A/T为中心的DNA的结合则仅受焓的驱动。
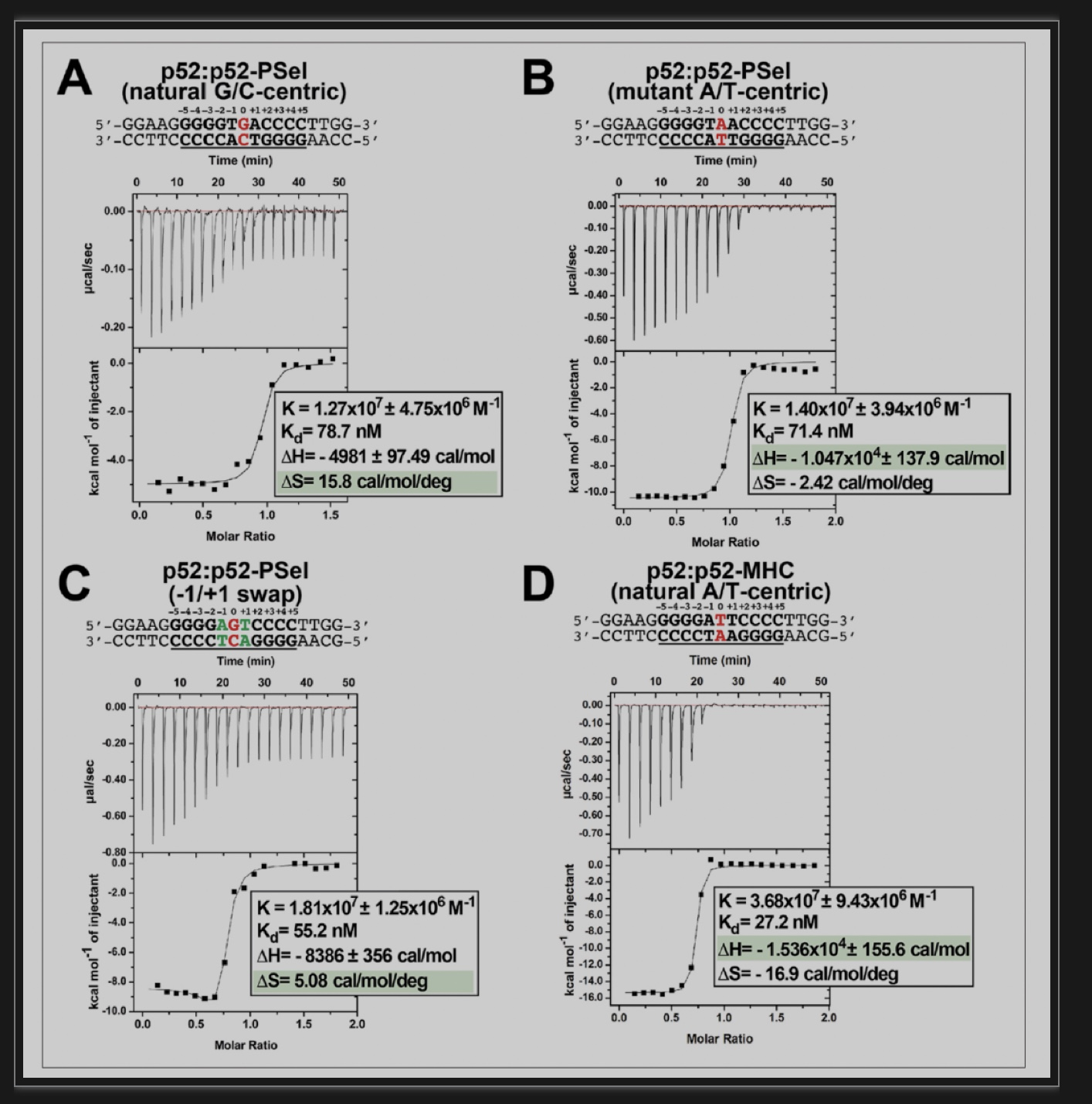
图4: (p52:p52)-DNA结合的热力学特征
(图源:Pan et al.,eLife,2023)
热力学结合机制并不能完全捕捉到由这些κB DNAs介导的不同的转录活性;因此,该团队更深入的研究了p52和κB DNA的结合动力学。有越来越多的证据表明,除了结合亲和力之外,动力学结合与解离速率(kon和koff)对各种细胞反应中蛋白质-配体相互作用的生理效果至关重要。结果表明,与其他两个变体和MHC-κB DNA相比,转录活性更强的天然的G/C为中心碱基PSel-κB DNA的结合显示出更高的结合率(kon)和解离率(koff)(图5)。而且,p52:p52同源二聚体与p52:p52:Bcl3复合物的结合动力学遵循同样的趋势:解离率(koff)越慢,(p52:p52)-DNA和(p52:p52:Bcl3)-DNA复合物的转录活性越低(图5K)。因此,转录活性可能与结合动力学有更密切的联系,而不是复合物的热力学稳定性。
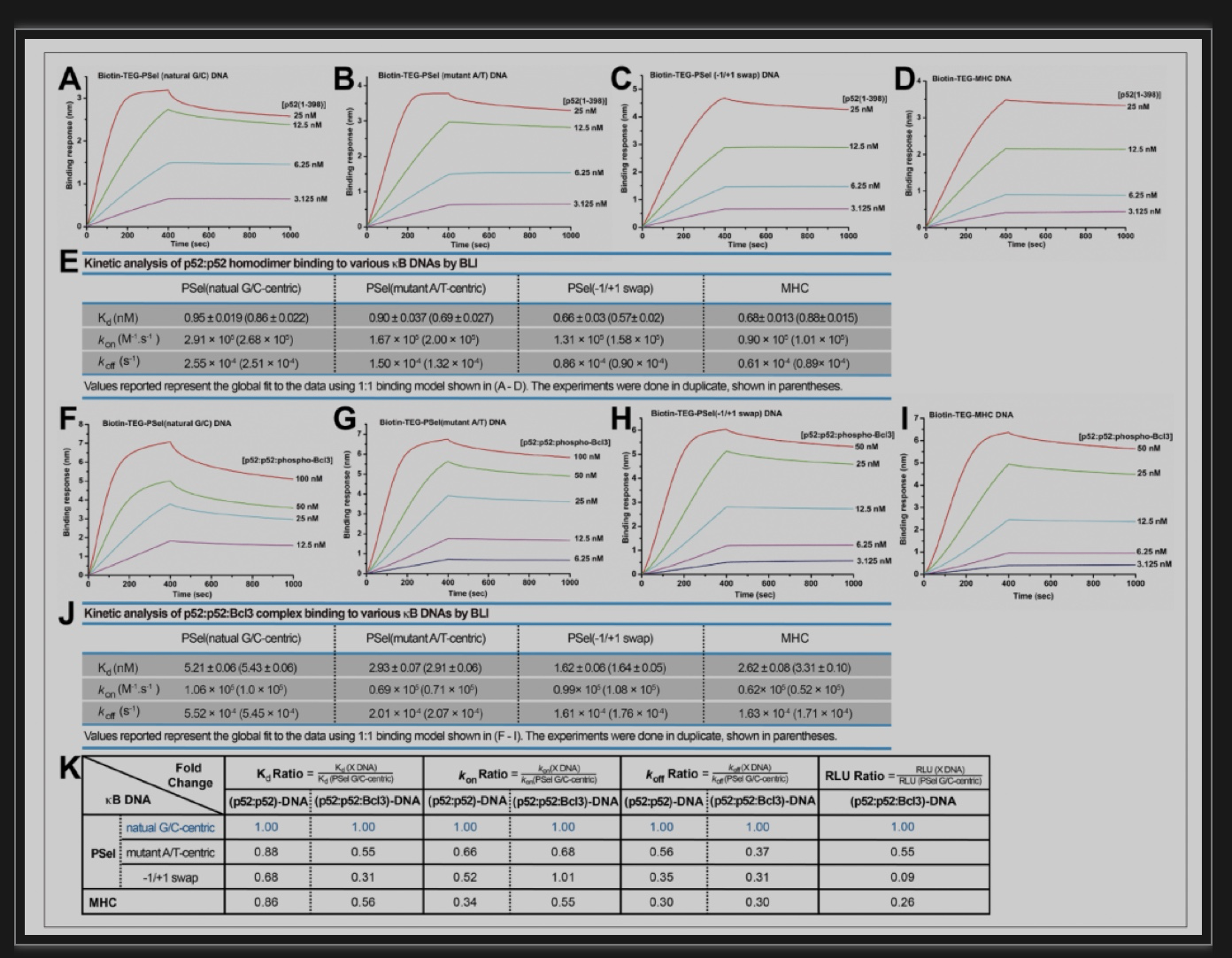
图5: p52同源二聚体与DNA结合的动力学特征
(图源:Pan et al.,eLife,2023)
