
II类内含子是一种可以自我剪接的逆转录元件,它们作为剪接体内含子和非长末端重复(LTR)逆转录元件的祖先,在塑造真核生物基因组中发挥着关键作用[1]。它们对于植物、真菌、酵母和许多细菌的基因表达非常重要[2,3]。II类内含子主要分为三类:IIA、IIB、IIC,它们的二级结构保守,三级结构相似并且都有核酶催化位点[4]。II类可以表达出特殊的逆转录酶,该类逆转录酶可以与产生它的内含子形成稳定的核糖核蛋白复合物,来帮助内含子的自我剪接[5]。同时所形成的核糖核蛋白复合物也可以作为逆转座子,攻击被剪切的外显子连接序列并通过酯交换反应,也就是反向剪接,整合进入基因组中。然而,现有的该类核糖核蛋白复合物结构并没有很好的阐明逆转录酶在核糖核蛋白的组装,底物DNA识别以及催化反应中的特殊机制,例如,为什么IIC内含子不仅识别DNA的序列而且还识别底物DNA的结构的机制也尚不清楚。这些问题妨碍了我们清晰的认识II类内含子的逆转座以及它在塑造基因组中的进化作用。
“Structures of a mobile intron retroelement poised to attack its structured DNA target”的研究。该研究通过冷冻电镜的方法,捕获了在逆转录状态下,IIC内含子与其表达的逆转录酶(MarathonRT),在有或者没有DNA底物情况下的形成的三元以及二元大分子复合物结构。该项研究阐述了逆转录元件的作用机制以及强调了II类内含子是遗传多样性的古老元素的假说。

该课题组利用冷冻电镜的方法,解析了一个2.8Å的高分辨率的IIC内含子和其表达的逆转录酶结合DNA底物的复合物结构(holoRNP)(图1)。该结构展示了内含子RNA和marathonRT以及紧密结合的底物DNA之间所形成的广泛的相互作用网络。尤其是独特的底物DNA,更是让该核糖核蛋白复合物区别于其他类内含子。
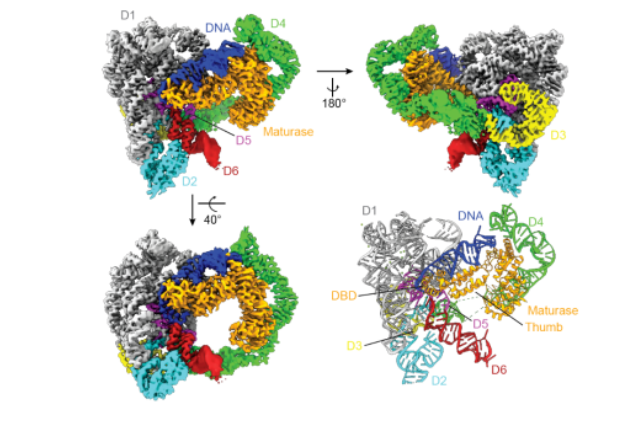
图1 IIC内含子转座元件的冷冻电镜结构
(图源:Chung & Xu, et al., Science, 2022)
该核糖核蛋白复合物结构捕获了其结合底物DNA但还没有进行反应时的状态(图2)。从解析的结构看,活性位点展示了内含子和它表达的逆转录酶之间相互依存的关系。在活性位点处,内含子RNA和它的底物DNA通过EBS-IBS(外显子-内含子)相互作用形成一段短的碱基配对(图2)。而这些短的碱基配对又受到逆转录酶支撑和固定,它们将逆转录酶的DBD(DNA结合结构域)的中间的α螺旋和thumb结构域的第三个α螺旋分别压向EBS1和EBS3,使其形成一个稳固的凹槽用来结合底物DNA。这些发现表明了逆转录元件的核心不仅仅只有RNA,它是一个由蛋白质和RNA相互作用而形成的活性位点。此外,该结构还展示了以前研究中没有提到过的逆转录酶的thumb和DBD结构域,帮助解释了RNA剪接以及内含子的整合依赖于逆转录酶是由于其紧密的相互作用,同时也强调了内含子RNA和它的辅因子蛋白在进化过程中是相辅相成的。
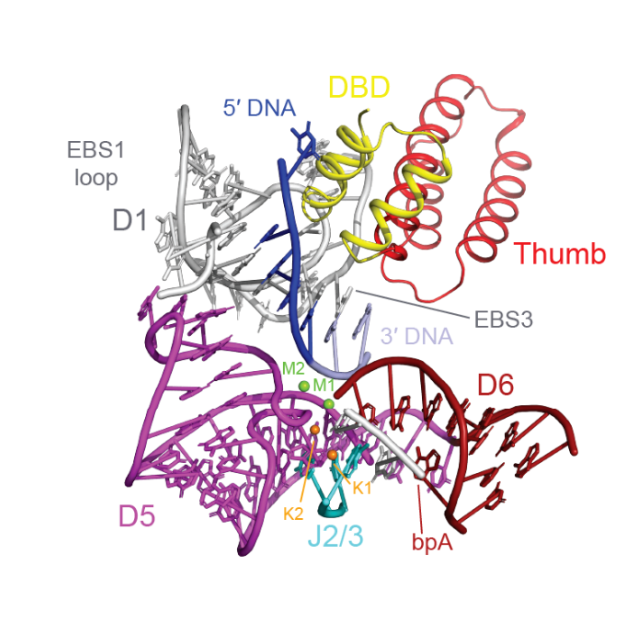
图2 内含子逆转录元件的活性中心
(图源:Chung & Xu, et al., Science, 2022)
此逆转座元件复合物具有一个较长的D4a结构域,它像一个从核酶结构域中心伸出的手臂,紧紧抱住逆转录酶(图3)。这个发现解释了之前报道中提到的D4a对于逆转录酶具有很强的亲和力,这种很强的亲和力正是由于逆转录酶和RNA之间具有广泛的相互作用。
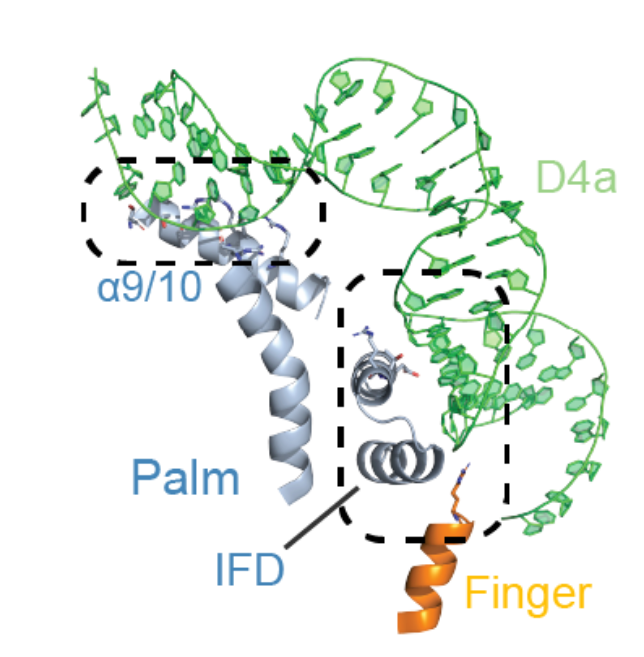
图3 蛋白与RNA的相互作用
(图源:Chung & Xu, et al., Science, 2022)
该结构还展示了RNA/蛋白复合物对于底物DNA识别的特殊策略。该种底物识别结合了结构特异性以及序列特异性。也就是说DNA本身独特的结构特征使其能够被该RNA/蛋白复合物结合。最典型的就是不同寻常的DNA结构,它包含一段由9个碱基对组成的沃森-克里克螺旋,紧接着是由G-A形成的非常规碱基配对和一个G-C碱基配对。这些配对延伸了该DNA的主干,使其满足IIC内含子结合进催化口袋的长度。接下来的DNA上的闭环结构中的碱基堆积在一起,像一个阶梯,连接着从核酶中心伸出的D4a螺旋(图4)。至此RNA/蛋白/DNA相互依偎在一起,形成一个稳定的复合物。
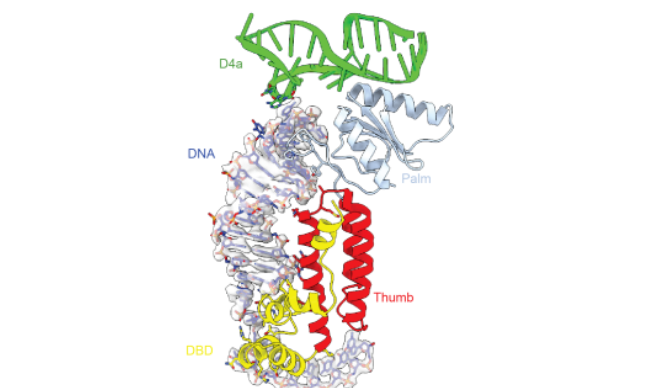
图4 特定结构的DNA底物与RNA/蛋白复合物之间的相互作用
(图源:Chung & Xu, et al., Science, 2022)
该研究组不仅解析了底物DNA与RNA/蛋白所形成的复合物,还捕获了尚未结合底物是时的RNA/蛋白符合物(apo)(图5)。通过对比两个复合物结构(RNA/蛋白/DNA和RNA/蛋白),该研究组发现,整个内含子与其逆转录酶所形成的复合物已经具备完美的底物结合口袋,随时准备与具有合适结构的DNA底物进入并发生反应。RNA蛋白复合物的催化中心的结构与结合底物后的结构保持了一致,逆转录酶也保留了原有的构象。但是在识别并结合底物DNA后,RNA蛋白复合物变得更加稳定。这和很多蛋白酶类似,进入结合口袋的小分子往往使局部结构更加稳定,从而把结合的小分子锁在结合口袋。
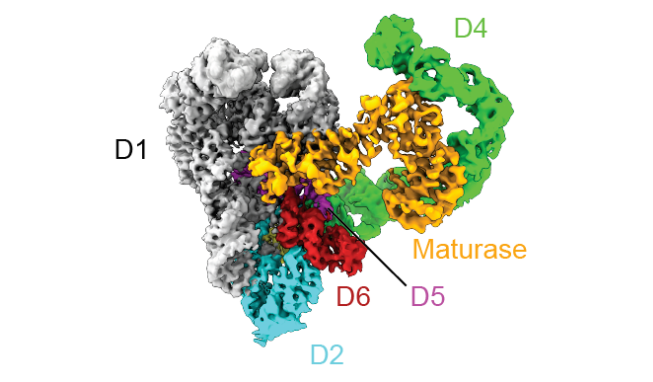
图5 未结合底物的RNA/蛋白复合物结构
(图源:Chung & Xu, et al., Science, 2022)
耶鲁大学Anna Marie Pyle研究组解析了高分辨率IIC内含子与其转录的逆转录酶结合以及未结合底物DNA的复合物结构。该项研究对于蛋白辅助的核酶催化提供了新的见解,同时拓展了对于RNA复合物识别DNA底物结构的认识,对于逆向剪接及逆转录具有启示意义。此外,该课题组对于II类内含子复合物的研究对于我们认识非长编码RNA转座子提供了一个新的思路,这也对相关疾病的研究提供了坚实的理论基础。但是该类内含子在剪接或者逆向剪接过程中还有一些其他的状态,未来工作中,研究者也将继续致力于捕获其他状态下复合物结构,通过这些结构希望可以全面阐述该类内含子是如何行使功能。
