
植物应对土壤多变的营养环境需整合和协调地上部和根系的养分感知信息,通过精细而复杂的信号转导机制,调控植物养分应答和生长发育进程。长距离信号转导机制的实现需经由维管系统进行信号分子的长距离运输 (故称长距离信号)。在众多矿质营养元素中,氮是限制植物生产力的主要元素之一。研究表明,蛋白质、小肽和 microRNAs 等多种分子均可作为长距离信号分子参与调控系统性氮信号转导。
在长期进化过程中,植物发展出复杂的机制来应对土壤环境变化,从而达到自身最优化生长和繁衍后代的目的。具体而言,就是植物体内存在一个稳健的营养信号调控网络,使其在异质的土壤环境中调控并维持植物自身生长。尤其在营养胁迫条件下,这个信号网络能够保证多种营养元素的协同利用和植物的适应性生长。
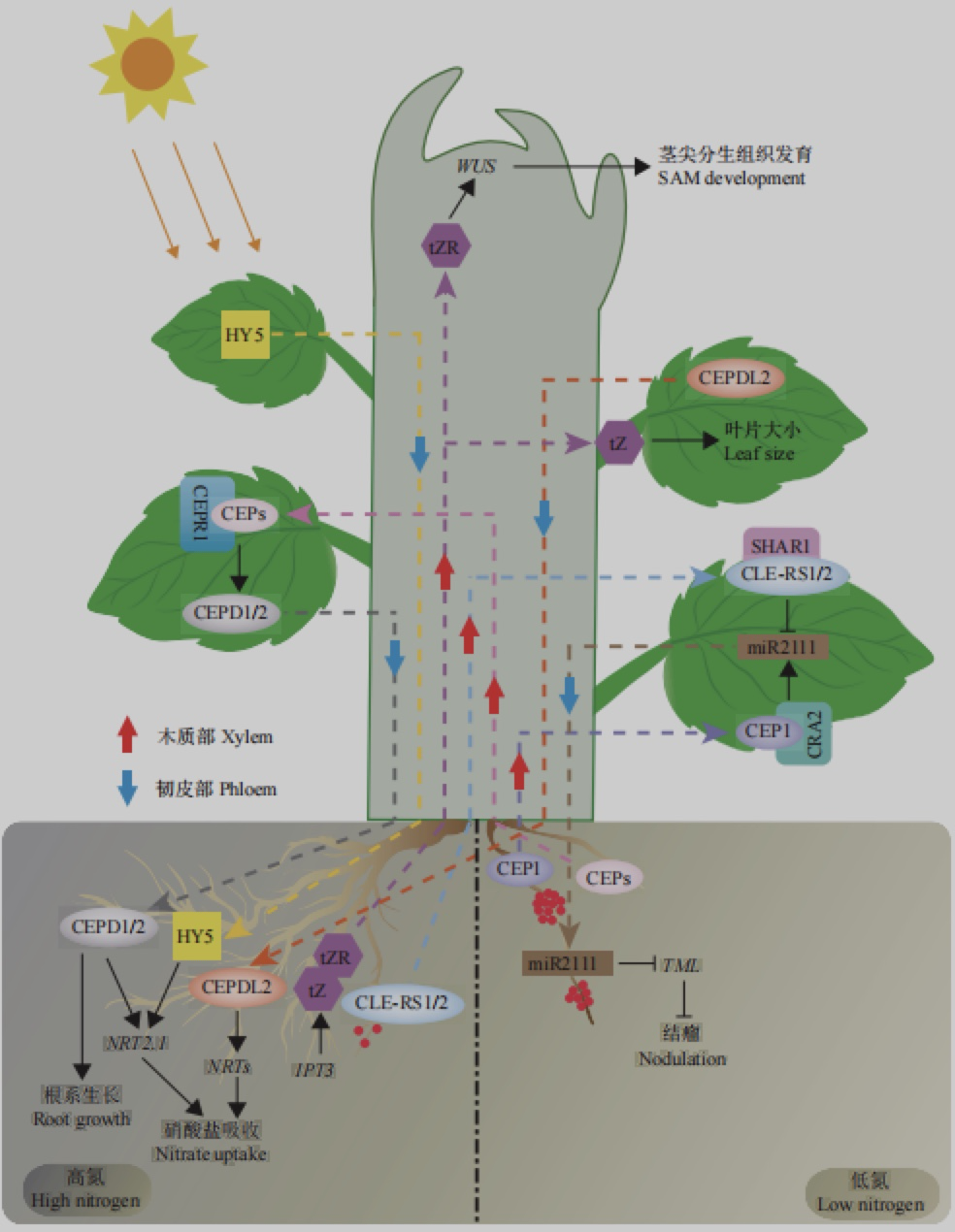
硝态氮 (NO3−) 和铵态氮 (NH4+) 是土壤中无机氮源的主要形式,在陆地土壤中,硝酸盐是多数植物获取氮源的主要形式。在硝酸盐分布均一的环境中,高浓度硝酸盐 (高硝) 抑制主根伸长和侧根发生,而低浓度硝酸盐 (低硝) 则促进主根伸长和侧根发生。在硝酸盐分布不均的自然环境下,植物根系通常表现出向营养性,即朝着高硝区域生长,同产生大量侧根来扩大根系与土壤接触面积;而低硝区域根系侧根伸长则被抑制。根系的氮营养向性反应则会在去除顶芽后消失,提示植物地上部分对根系的可塑性调控具有重要作用,亦即植物存在长距离营养信号系统调节植物根系的可塑性。
1.小肽介导的系统性信号调控植物整体的氮吸收利用
分根试验表明,低硝不仅能促进高硝侧的植物侧根增多,还能通过诱导高硝侧根系中 NRT2.1 的表达来促进高硝侧根系对外界 NO3−的吸收。进一步研究发现,低硝会诱导根系产生 15 个氨基酸的小肽 CEPs ,并通过木质部长距离运输至地上部,而后被 LRR 受体激酶CEPR1识别,CEP-CEPR 信号增强并诱导合成 CEPD1 和 CEPD2 ,它们通过韧皮部由地上部长距离运输至高硝一侧的根系中,诱导 NRT2.1 基因的表达,提高根系吸收硝酸盐的效率。除调控硝酸盐吸收相关基因转录外,CEP-CEPR 信号通路还可抑制生长素由地上部向根的转运,进而调控根系构型和侧根夹角,增大根系面积。同时,低硝通过诱导CLE小肽家族的基因 CLE1/3/4/7 的转录,抑制侧根原基生长。组织定位分析表明,CLE1/3/4/7 可能由中柱细胞分泌,并被转运至韧皮部伴胞细胞中,从而被定位于韧皮部的受体蛋白 CLV1识别后,产生系统性氮信号,负调控根系发育。
植物不仅能通过根感知外界土壤中的氮营养状态,也能通过地上部感知整体氮营养状态并产生长距离信号调控根系对氮的吸收利用。植物地上部缺氮会诱导叶片产 生 CEPD1/ 2 的同源蛋 白CEPDL2 ,CEPDL2 通过韧皮部长距离运输迁移到根中,促进硝酸盐转运相关基因NRT2.1、NRT1.5、NRT2.2、NRT2.4 和 NRT3.1 的转录,提高根系氮吸收能力,从而实现植株整体发育的协调。因此,小肽在协调植物整体氮吸收和利用中发挥重要作用。
2. 小肽介导的系统性信号调控豆科植物结瘤固氮
豆科植物根系不仅可以通过吸收土壤中的硝酸盐和铵盐获取氮素,还可通过与根瘤菌共生进行生物固氮。然而,共生固氮高度耗能,豆科植物会根据外界氮源供应情况调控结瘤,即在缺氮环境中,豆科植物通过消耗碳源形成根瘤进行生物固氮;在氮充足的环境中,过度结瘤会损失碳源,抑制宿主生长,因此,豆科植物启动根瘤自主调控系统(AON),抑制根瘤形成,避免宿主植物过度结瘤而浪费碳源。研究表明,一些 miRNA、小肽和蛋白质等信号分子参与了由硝酸盐介导的根系结瘤过程。
低硝会诱导蒺藜苜蓿、百脉根和大豆等豆科植物根系产生 CEP1 小肽,并通过木质部长距离转移到地上部与受体 CRA2 结合,促进 miR2111 前体转录,此后 miR2111 通过韧皮部长距离运输至根部直接抑制结瘤负调控因子 E3 泛素连接酶基因 TML 表达,促进结瘤。在高硝条件下,根瘤菌侵染豆科植物后会诱导 CLEs 基因的表达,CLEs 通过木质部长距离转运至地上部,从而激活 AON 途径并抑制结瘤。在高硝条件下,百脉根中 NIN转录因子会促进根中 CLE-RS1/2 小肽合成,而后经木质部长距离运输至地上部与受体激酶 LjHAR1 结合,进而抑制根瘤形成。研究表明,CLE-RS1/2-LjHAR1 复合体可能通过以下2 个途径影响结瘤:一方面抑制叶片中 miR2111合成,导致由地上部转运到根中的 miR2111 减少,从而减弱对 TML 基因的转录水平抑制,阻碍根瘤菌侵染;另一方面,CLE-RS1/2-LjHAR1 复合体可能通过激活细胞分裂素合成基 因 IPT3的表达促进细胞分裂素合成,细胞分裂素转运至根部抑制结瘤。但是,细胞分裂素抑制结瘤的分子机制仍不清楚。同样,在蒺藜苜蓿中,高硝促使 NIN-like 家族成员 MtNLP1 激活 MtCLE35 的转录,翻译产生的 MtCLE35 被转运至地上部与 MtSUNN 受体蛋白结合,通过抑制 miR2111表达影响根系结瘤。综上所述,高氮通过NIN-CLE-AON 途径抑制结瘤在多数豆科植物中是保守的。
3. 细胞分裂素介导的长距离氮信号调控植物生长发育
充足的硝酸盐能促进植物地上部的快速生长,然而根中的氮信号传递至地上部分并促进地上部生长发育的分子机制并不清楚。研究发现,硝酸盐处理后,木质部中细胞分裂素前体 tZR浓度升高,暗示在根部硝酸盐信号可能会转换成细胞分裂素信号。进一步研究发现,硝酸盐可以诱导拟南芥细胞分裂素合成基因 IPT3 的表达,导致根系中细胞分裂素前体 tZR 和有活性的细胞分裂素 tZ 增加,而后 tZR 和 tZ 通过ABC 类转运蛋白 AtABCG14 长距离运输至地上部分调控生长发育。被转运至地上部的细胞分裂素可在不同的部位发挥功能:tZ 主要调控叶片大小,而 t ZR 通过影响 WUSCHEL ( WUS ) 和CLAVATA3 (CLV3) 的表达调控茎尖分生组织的活性,进而调控植物地上部分的发育。此外,细胞分裂素到达地上部分后,会促进地上部谷氨酸和谷氨酰胺合成,这些氨基酸可能作为系统性信号从地上部分转移到根调控硝酸盐的吸收。
在水稻中,硝酸盐和铵盐的代谢产物是谷氨酰胺,硝酸盐、铵盐和谷氨酰胺均能诱导根中 OsIPT4、OsIPT5、OsIPT7 和 OsIPT8 的表达,但谷氨酰胺合成酶抑制剂 L−蛋氨酸亚砜酰亚胺 处理则能阻断硝酸盐和铵盐对OsIPTs 的诱导,说明是谷氨酰胺直接诱导根中OsIPTs 的表达。研究还发现,OsIPT4 是根中细胞分裂素合成的主要基因,其 RNAi 株系的木质部细胞分裂素含量减少,地上部各器官表现缺氮表型。上述结果表明在单双子叶植物中,细胞分裂素介导的长距离氮信号是高度保守的。
4. 光信号及光合产物对长距离氮信号的影响
植物对许多矿质元素的吸收都受到光信号和光合产物的影响,光合产物能够促进侧根生长,并诱导硝酸盐转运蛋白基因的表达促进硝酸盐吸收。进一步研究发现,光合代谢产物蔗糖可以诱导多个 AtCEP 基因 (AtCEP5~AtCEP9) 的表达,AtCEPs 被 AtCEPR1 识别并通过未知的机制限制侧根生长,并且这个过程需要依赖于 AtCEPR1 蛋白活性,上述结果表明,在拟南芥中CEP-CEPR1 模块能够整合碳信号和氮信号并作为负反馈环调控根系的适度生长。最新鉴定到植物感知光合产物匮乏调控硝态氮响应的关键组分 SnRK1。SnRK1 是一个保守的能量感受器,其催化亚基 KIN10 突变抑制了由光合产物匮乏所致的硝酸盐应答反应,暗示 KIN10 是光合作用与硝酸盐信号交互调控的重要枢纽。进一步研究发现,光合产物匮乏促使 KIN10 磷酸化 NLP7 并使其滞留在细胞质中被降解,进而抑制下游的硝酸盐应答反应。但 KIN10 如何感知光合产物匮乏并使 NLP7 降解的机制并不清楚,光合产物如何调控氮吸收利用相关基因表达的分子机制仍然未知。
在拟南芥中,光信号通路中的核心转录因子HY5 是碱性亮氨酸家族成员,能接收来自不同的光感受器捕获的光信号,包括光敏色素、隐花色素、紫外线 B ,并将信号传递给下游信号元件,进而调控光形态建成、根系发育、非生物胁迫响应、养分吸收等植物生长发育过程。研究发现,HY5 蛋白作为一个长距离信号分子还参与植物地上部和根系间的远程交流,协调生长发育和碳/氮平衡。拟南芥 HY5 在地上部促进 TPS1、SWEETs 等基因的表达,增强了碳同化和碳转运,并在光的作用下通过韧皮部转移到根,激活根中 HY5 和 NRT2.1 的转录,调节根的生长和硝酸盐的吸收。因此,HY5 作为一个长距离信号分子协调了受光响应的碳固定和氮吸收,使植物地上部和根实现协同生长。
