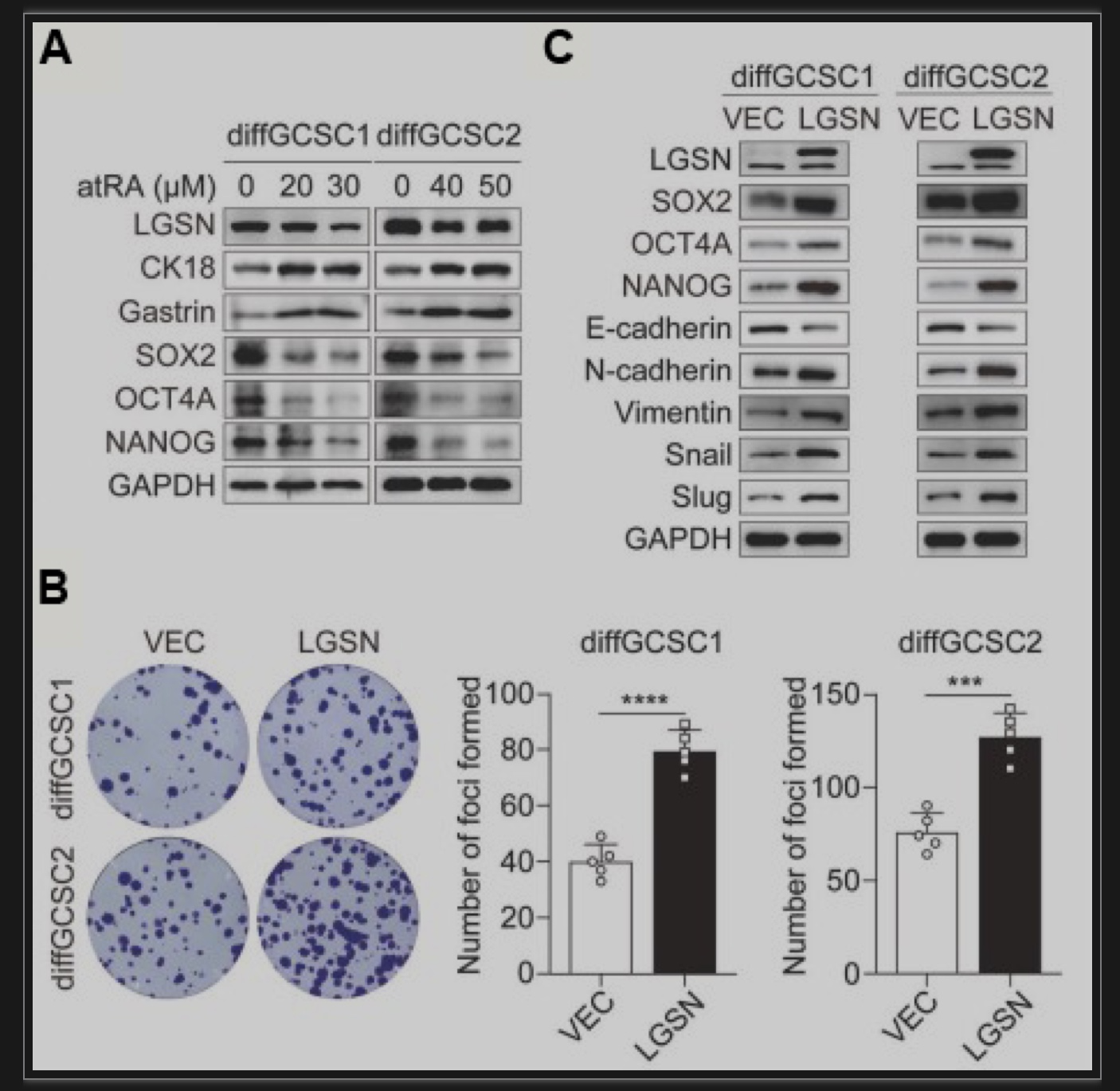
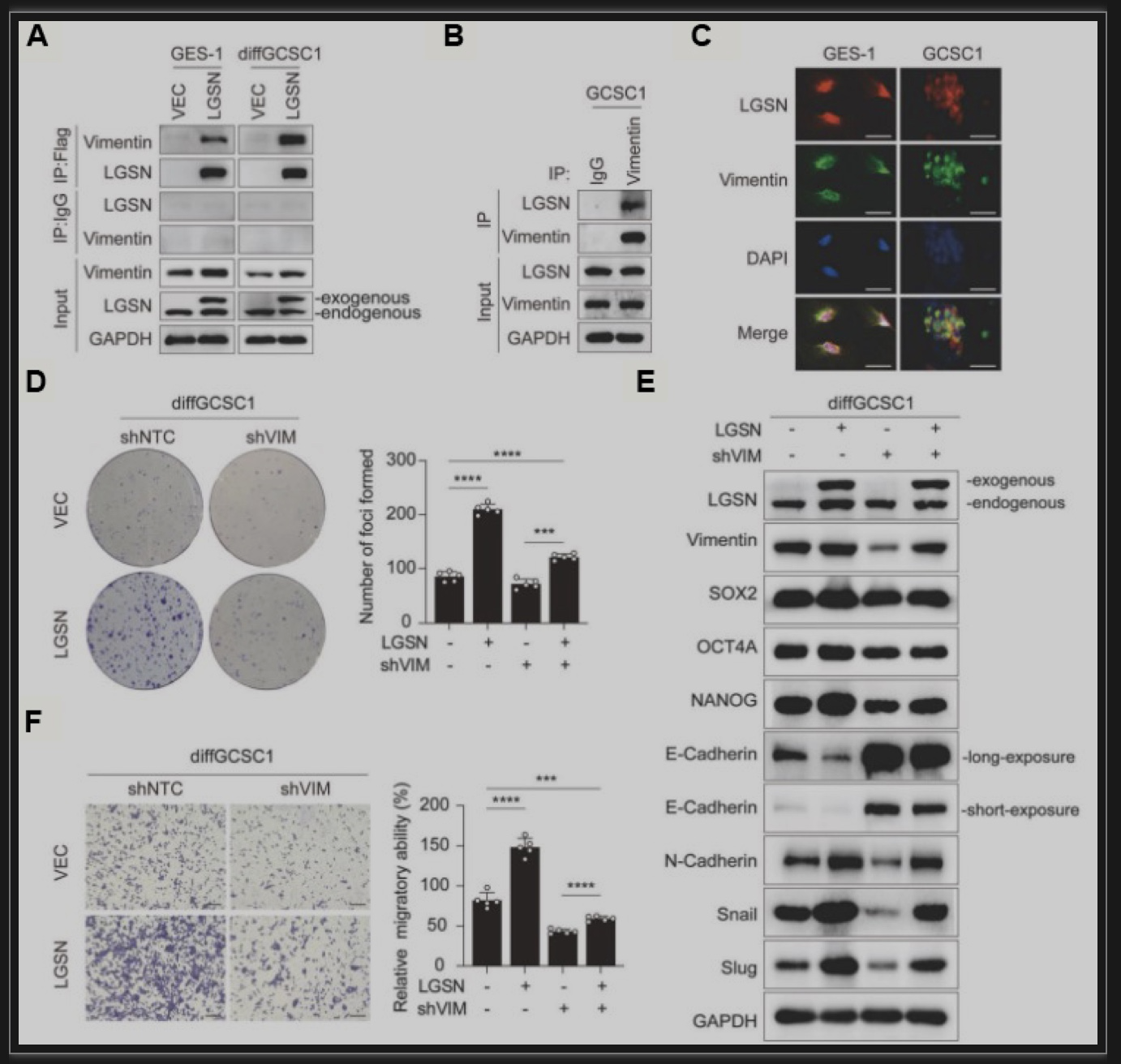
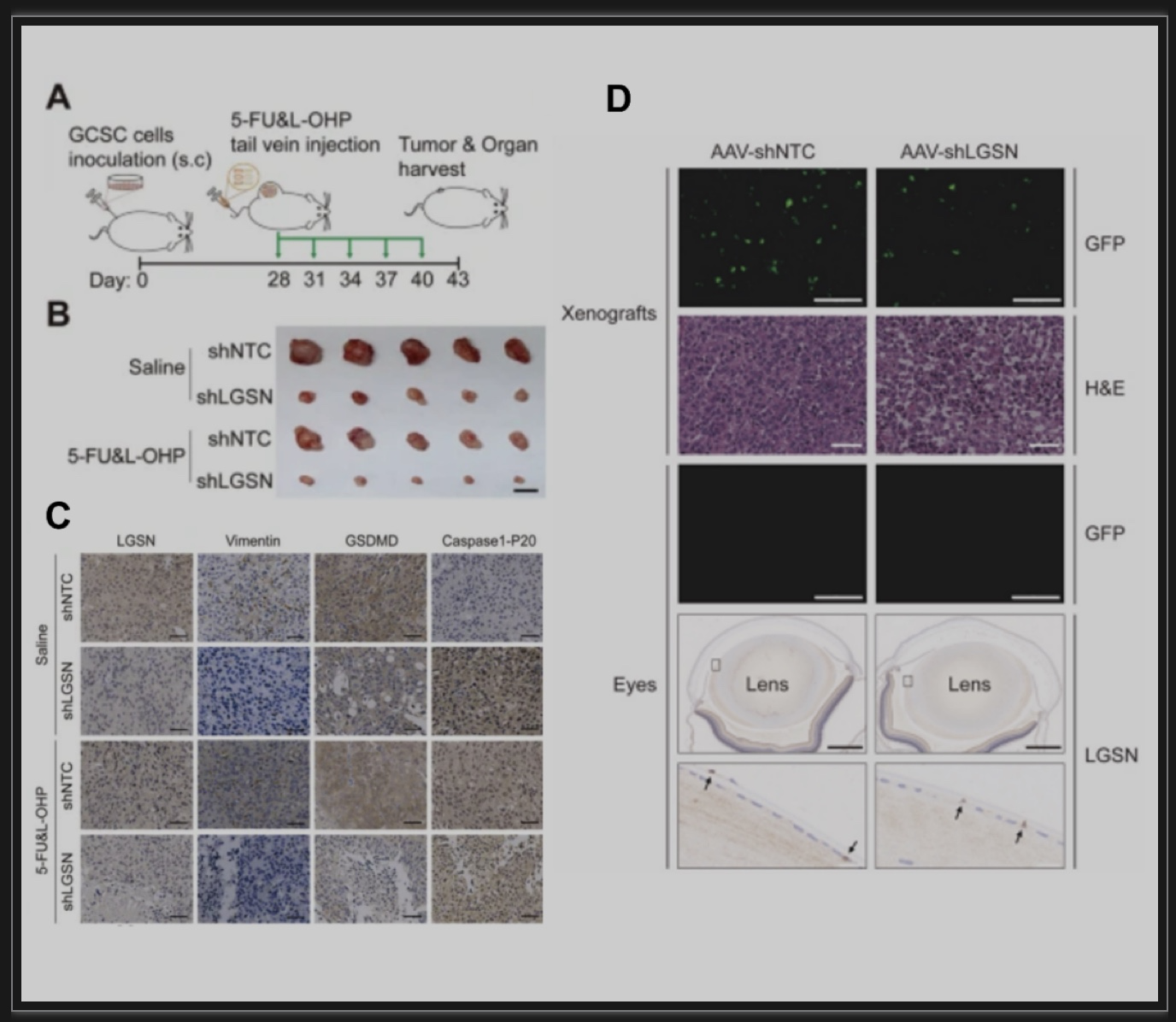
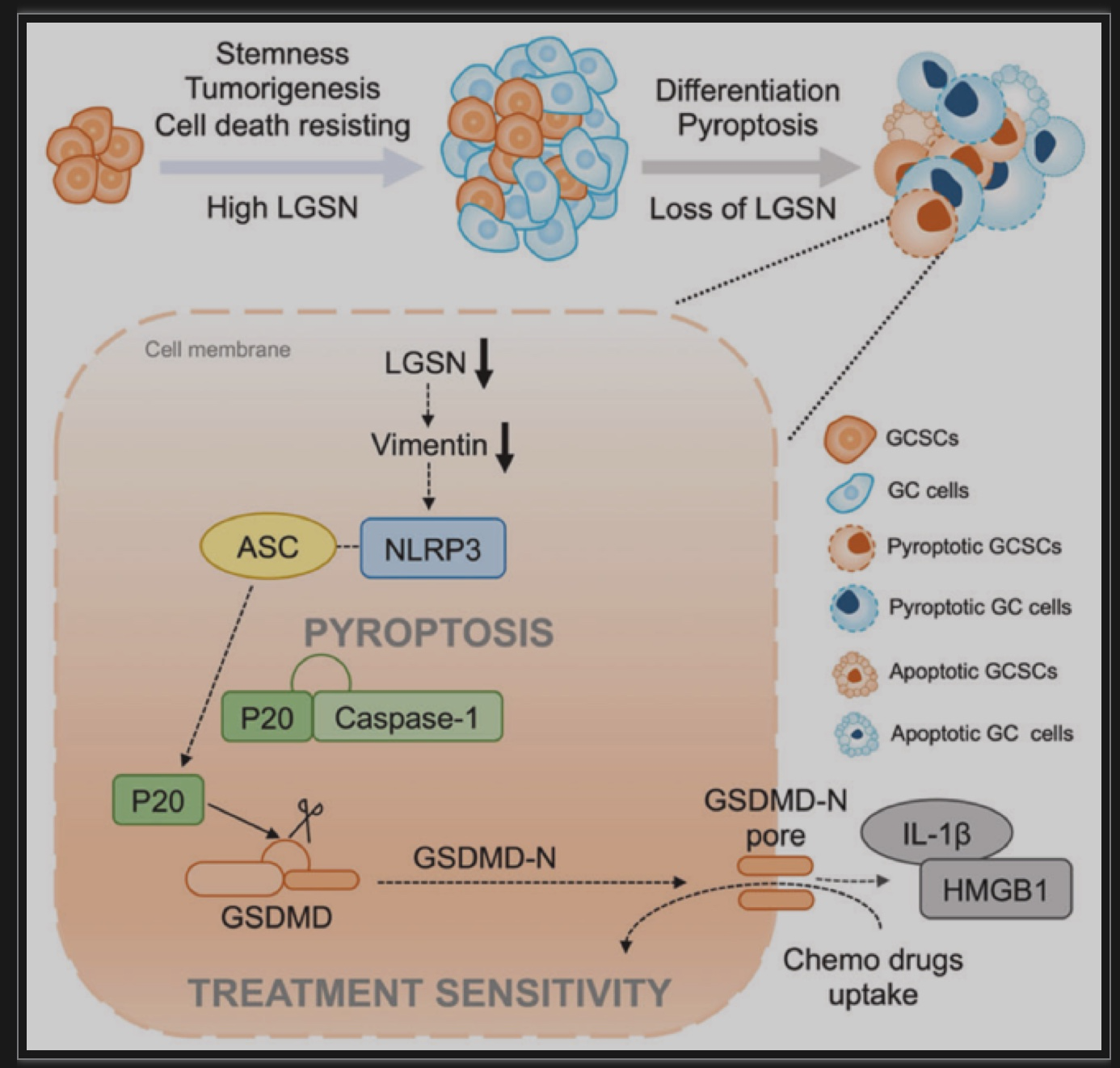
胃癌是世界范围内发病率和死亡率最高的恶性肿瘤之一。正常的成体干细胞可以通过致瘤信号刺激转化为肿瘤干细胞(Cancer Stem Cells, CSCs)并分化为肿瘤细胞。由于胃上皮成体干细胞具有多向分化潜能,其癌前突变可能是胃癌发生的直接原因[1, 2]。虽然大多数癌症的化疗耐药、转移和复发都归因于CSCs的持续再生,但对CSCs逃避死亡的关键特性和确切机制以及潜在的可干预靶点知之甚少[3]。揭示了靶向LGSN通过诱导焦亡恢复胃癌干细胞化疗敏感性的分子机制。该研究通过RNA-seq比较胃癌患者来源的CD44+CD54+胃癌干细胞在贴壁(分化状态)及成球(未分化状态)培养时的差异基因表达,发现LGSN在胃癌干细胞成球时显著上调,同时伴随着干性相关基因的上调,其中包括经典的干性相关因子SOX2、OCT4A以及NANOG。Lengsin(LGSN)是晶状体中一个关键的中间丝终末分化相关调节因子。迄今为止,LGSN的主要研究性工作主要集中在晶状体和白内障疾病上[4, 5],而在肿瘤乃至胃癌的研究上仍存在很大程度上的空白。进一步利用TCGA胃癌数据集进行基因集富集分析,发现LGSN与胃癌进展和干细胞亚群维持显著相关。以上结果提示了LGSN与胃癌干性的关联(图1A-E)。图1 LGSN与胃癌细胞干性显著相关(图源:Li YT et al.,Cell Death Dis, 2023)使用分化诱导剂atRA对胃癌干细胞进行处理后发现,LGSN表达降低并伴随着干性相关因子SOX2、OCT4A和NANOG的下调以及胃细胞分化标志物CK18和Gastrin的上调(图2A)。过表达LGSN能够促进胃癌干细胞的干性、增殖能力(图2B)、以及上皮-间质转化(图2C),表明LGSN能够显著的促进胃癌干细胞的恶性生物学行为。图2 LGSN增强胃癌干细胞的干性,促进增殖和上皮-间质转化(图源:Li YT et al., Cell Death Dis, 2023)为了探索LGSN的促癌机制,对LGSN蛋白进行免疫共沉淀并进行质谱分析寻找其相互作用的蛋白。作者鉴定出Vimentin为LGSN的互作蛋白并证明了Vimentin介导了LGSN调控胃癌干细胞的促癌特性(图3A-F)。图3 LGSN与Vimentin的相互作用(图源:Li YT et al.,Cell Death Dis, 2023)为进一步了解LGSN如何影响胃癌干细胞的自我更新和致瘤能力,作者在胃癌干细胞敲低LGSN后,发现胃癌干细胞的死亡显著增加(图4A)。死亡的细胞形态发生明显的肿胀,质膜上出现特征性的大泡状突起(图4B),表明细胞经历了以焦亡为特征的形态学变化[6]。免疫印迹试验对细胞焦亡相关蛋白NLRP3、ASC、pro-caspase1、caspase1-P20以及GSDMD-N检测后发现,LGSN敲低激活了焦亡信号通路以及促进了随后从细胞膜释放的IL-1β、HMGB1和LDH(图4C,D)。此外,随着LGSN的敲低,Vimentin蛋白的表达也逐渐降低(图4E)。这些结果说明LGSN能够促进Vimentin蛋白的表达以及抑制胃癌干细胞的焦亡信号通路的激活。图4 LGSN敲低激活焦亡相关信号通路(图源:Li YT et al., Cell Death Dis, 2023)作者在裸鼠皮下成瘤模型中进一步验证了LGSN敲低增强了胃癌干细胞对化疗药物的敏感性(图5A-C)。此外,为了评估LGSN作为胃癌靶点治疗的安全性,作者通过尾静脉注射GFP标记的腺相关病毒AAV-shLGSN和AAV-shNTC,对胃癌干细胞荷瘤小鼠进行了全身治疗。结果表明,与AAV-shNTC治疗相比,AAV-shLGSN治疗可有效抑制胃癌干细胞异种移植小鼠的成瘤性,并且不会影响LGSN在晶状体中的表达,证明了LGSN作为胃癌治疗靶点的有效性和安全性(图5D)。图5 LGSN敲低增加胃癌干细胞化疗敏感性,且腺相关病毒靶向敲低LGSN不影响晶状体中LGSN的表达。(图源:Li YT et al.,Cell Death Dis, 2023)图6 LGSN介导的胃癌干细胞焦亡抵抗机制图(图源:Li YT et al.,Cell Death Dis, 2023)文章结论与讨论,启发与展望胃癌微环境表现出高度的异质性,其包含了不同谱系和分化水平的细胞。胃癌干细胞在启动和维持肿瘤中发挥着关键作用,胃癌细胞的去分化可以促进胃癌干细胞的出现。然而,胃癌干细胞产生及维持的决定因素至今仍未完全阐明。本研究中,作者描述了胃癌干细胞中LGSN通过与Vimentin相互结合,从而抑制下游焦亡信号通路的激活,维持细胞膜完整性,降低化疗药物的摄取,最终造成胃癌化疗抵抗的分子机制。该研究通过揭示LGSN在胃癌干细胞中的功能与机制,不仅有助于更好地了解LGSN在除了晶状体以及眼部疾病之外的生理和病理作用,而且还为胃癌化疗耐药提供新的干预靶点。