18101298214
公司首页
实验手册
小分子化合物
细胞培养和检测
定制服务
文献资料
联系我们
订购指南
公司首页
实验手册
小分子化合物
细胞培养和检测
定制服务
文献资料
联系我们
订购指南
首页
实验手册
发现新的减数分裂重组交换蛋白C12ORF40/REDIC1
发布日期:2023/8/31 13:50:00
不孕不育
困扰高达17.5%的育龄夫妇[1],其中大多数都是由于配子(精子和卵子总称)发生和成熟障碍导致的。减数分裂是配子发生的必需环节,在减数分裂I前期,同源染色体首先进行相互识别和配对,再通过重组形成交换才能在同源染色体之间建立起物理连接,从而确保它们能够正确地排列到减数分裂中期I的纺锤体赤道板以及后期的分离和配子发生减数分裂的正常进行[2,3]。因此,重组交换是减数分裂正常发生和配子形成的关键。减数分裂重组起始于程序性DNA双链断裂(Double strand break,DSB)的产生和以同源染色体上的DNA为模版进行的修复[4]。值得注意的是,人或小鼠的每个精母细胞会产生250-350个DSBs,但其中只有约10%经过修复而形成交换(Crossover,CO)。因此,重组交换受到严格的调控[5-8]。
虽然已有多种蛋白被报道参与重组交换,但交换形成的调控机制仍不太清楚。
该研究首次报道了一个新的减数分裂重组修复关键蛋白C12ORF40/REDIC1,该蛋白通过稳定重组中间体进而促进减数分裂交换形成;并证实了C12ORF40/REDIC1的功能丧失突变会导致人类精子发生减数分裂重组减少和无精子症发生。
首先,该团队通过全外显子组测序在两例无血缘关系的散发
非梗阻性无精子症
(Non-obstructive azoospermia,
NOA
)患者中筛选到一个功能未知基因
C12orf40
的纯合移码突变(c.232_233insTT, p.Met78Ilefs*2),并根据该突变导致人和小鼠第一类重组交换减少的表型将
C12orf40
命名为REDIC1 (Reduction in class I crossover)。随后,作者以小鼠为模型,对
C12orf40
在小鼠中的同源基因
CN725425
/
Redic1
的表达、定位及功能进行了深入研究。
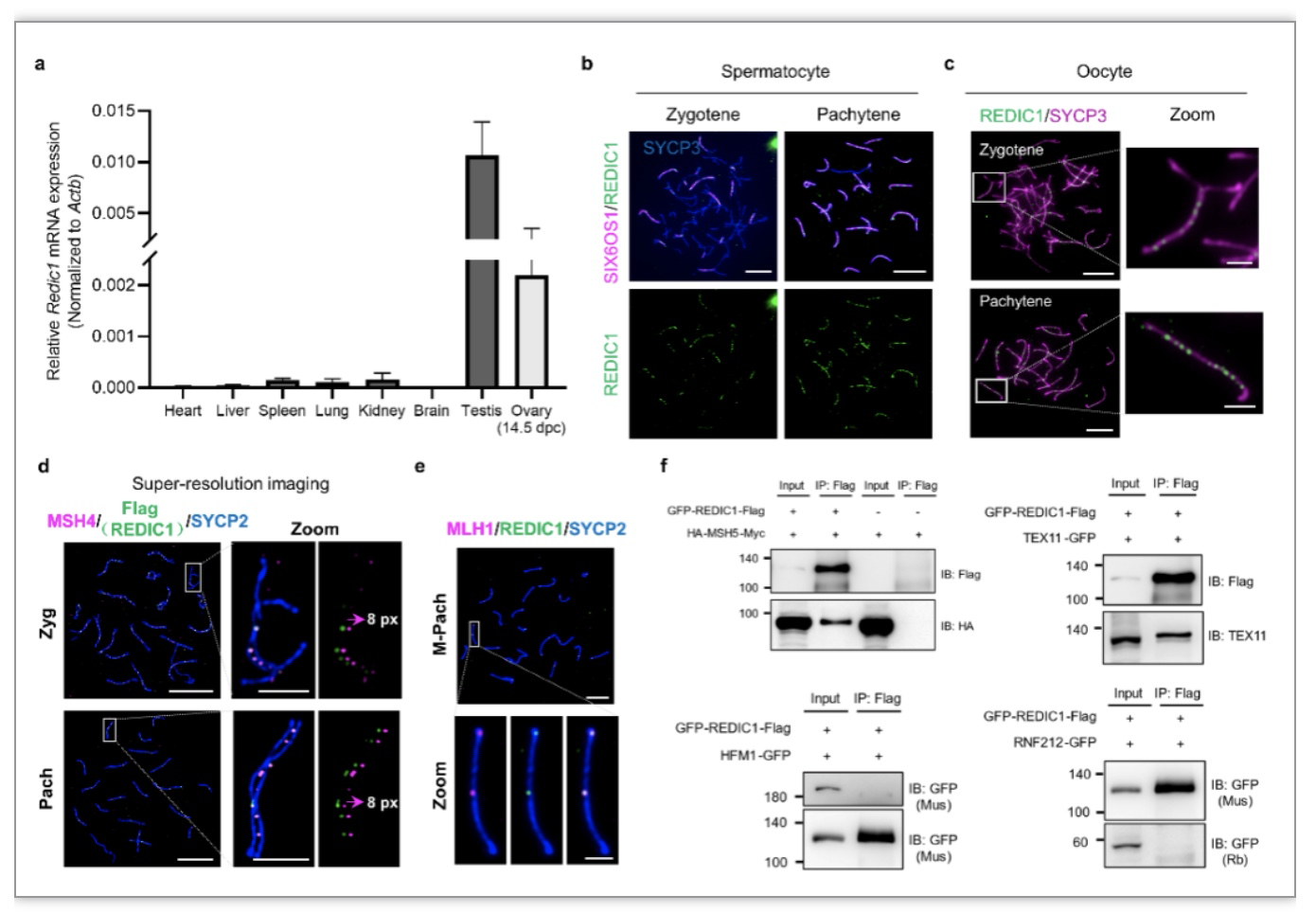
Redic1
在成年小鼠睾丸及胚胎期的卵巢组织中高表达,提示它可能在减数分裂前期发挥作用。免疫荧光染色结果显示REDIC1蛋白从偶线期开始呈点状信号定位在精母细胞和卵母细胞中同源染色体的配对区域,随后数目逐渐增多,在晚偶线期达到峰值,随后数目又逐渐减少,在中晚粗线期细胞中的数目与CO数目接近,进入双线期后消失,提示REDIC1可能是一种减数分裂重组蛋白。利用
Six6os1
-/-
、
Top6bl
-/-
和
Dmc1
-/-
小鼠精母细胞进行的荧光免疫染色显示,REDIC1的定位依赖程序性DSBs的产生以及链入侵,但不依赖同源染色体联会,这种定位模式与ZMM蛋白类似。随后,作者通过共染及超高分辨显微成像发现REDIC1与ZMM蛋白之一的MSH4共定位,并在中晚粗线期精母细胞中与晚期重组结蛋白MLH1共定位。免疫共沉淀实验进一步证实了REDIC1与ZMM蛋白MSH5(MSH4的互作蛋白)和TEX11直接互作(图1)。
这些结果表明REDIC1是一个新的减数分裂重组蛋白。
图1. REDIC1的表达及定位分析
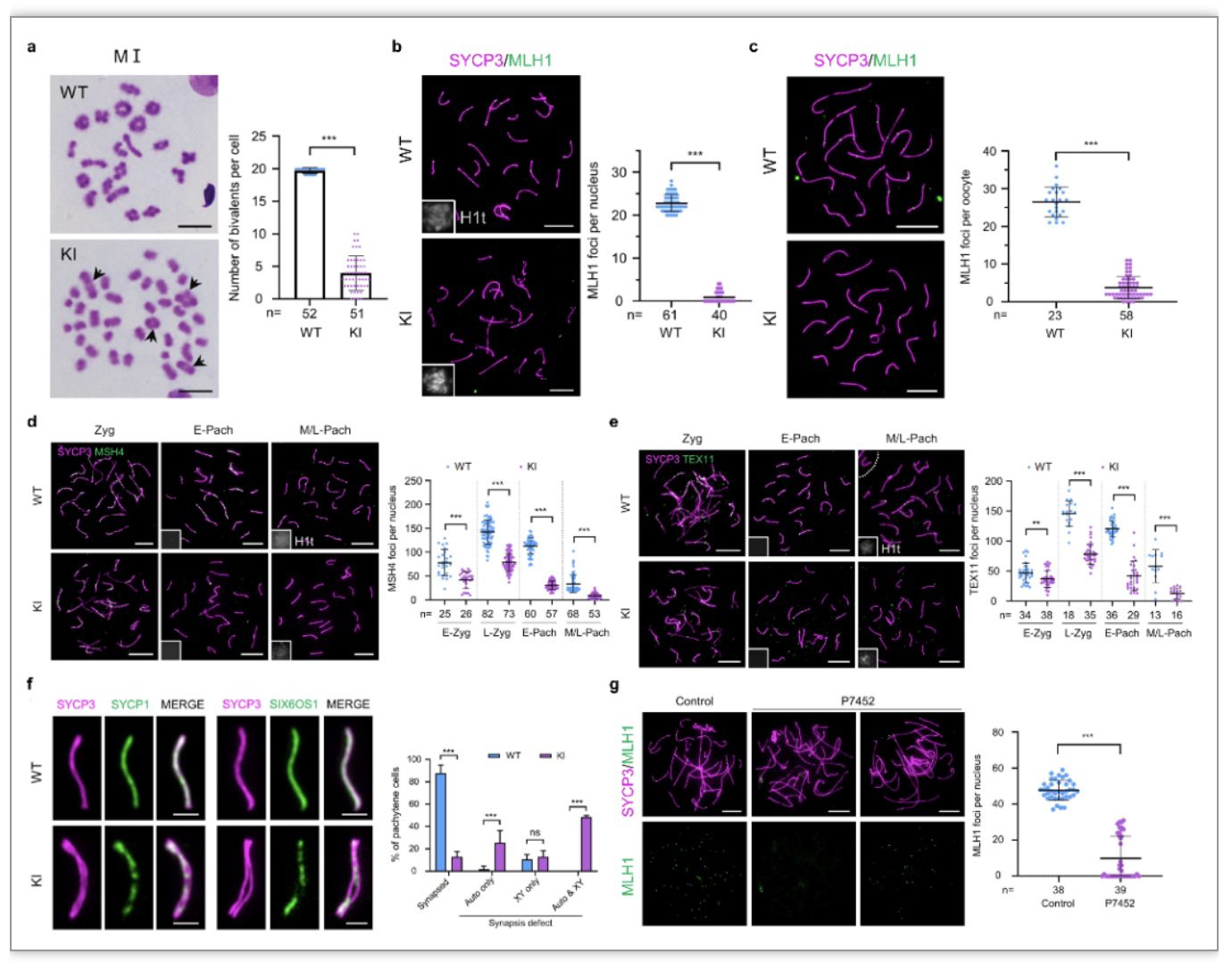
接着,为了验证在患者中发现的c.232_233insTT突变的致病性,该团队制备了模拟患者突变的
Redic1
基因突变敲入(Knock-in,KI)小鼠模型(
Redic1
-KI),并通过睾丸组织学分析,发现该突变的确会导致小鼠精子发生停滞于减数分裂中期Ⅰ,不能产生精子。进一步分析发现,
Redic1
-KI小鼠的中期Ⅰ精母细胞中的二价体数目显著减少(野生型小鼠的每个细胞有20个二价体,突变小鼠每个细胞中平均只有4个)。与此一致的是,
Redic1
-KI小鼠每个中粗线期精母细胞中标志重组交换的MLH1数目只有约1个,远远少于野生型小鼠的约23个,这表明REDIC1对于CO形成至关重要。此外,
作者发现
Redic1
突变也会导致雌性小鼠的卵母细胞CO数目减少、卵泡形成和发育严重受损。
为了明确
Redic1
-KI小鼠CO数目减少的原因,作者对不同发育阶段精母细胞中多种ZMM蛋白的信号点数目进行了统计,发现
Redic1
-KI小鼠的MSH4和TEX11数目均较野生型小鼠的少。这提示
Redic1
突变可能通过导致重组中间体减少而诱发CO数目减少,同时也提示REDIC1能够促进MSH4和TEX11在染色体轴上的积累(图2)。此外,作者还发现
Redic1
突变会导致一些染色体出现不完全联会。更重要的是,
CO数目减少以及联会缺陷的表型在患者精母细胞中得到了重现,进一步证实了
C12orf40
c.232_233insTT纯合移码突变确实是患者的致病突变。
图2.
Redic1
突变导致联会和交叉形成异常
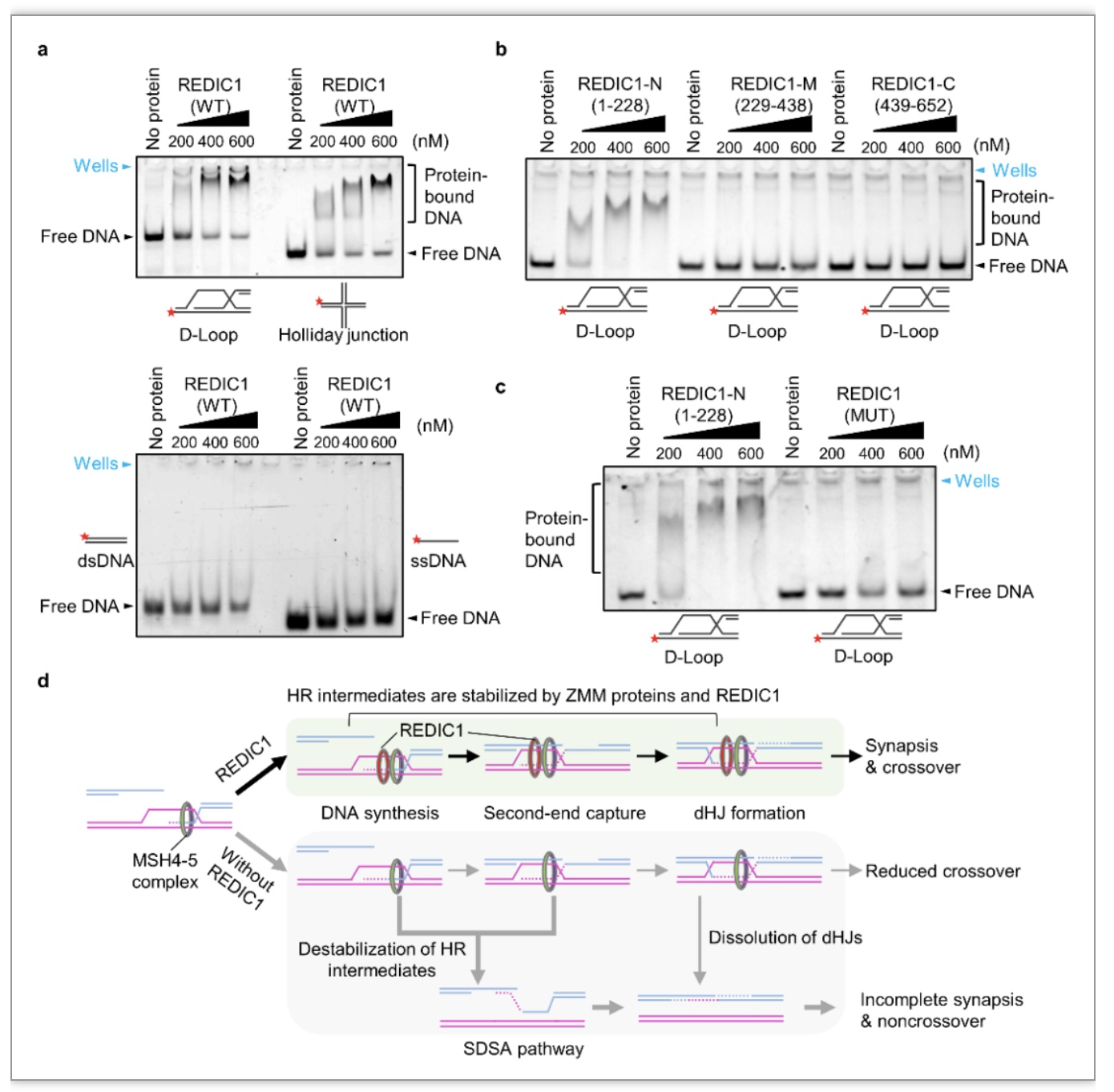
最后,为了进一步揭示REDIC1在减数分裂重组中的作用,该团队表达和纯化了REDIC1蛋白,并在体外将其分别与单链DNA(single-stranded DNA,ssDNA)、双链DNA(double-stranded DNA,dsDNA)、含有D-loop结构和Holliday junction(HJ)结构的DNA孵育,并进行了EMSA实验。结果显示REDIC1全长蛋白对含有D-loop和HJ的DNA底物的结合远强于对ssDNA和dsDNA的结合。随后,
该团队发现REDIC1的N端220多个氨基酸负责其与含有D-loop结构的DNA底物的结合,而REDIC1突变蛋白丧失了与DNA底物的结合
(图3)
。
图3. REDIC1促进减数分裂交叉重组
文章结论与讨论,启发与展望
综上,本研究首次报道了
C12orf40
/
REDIC1
纯合移码突变会导致人类非梗阻性无精子症,为相关不育患者的遗传咨询和临床诊疗提供了分子靶标。同时,本研究从多个角度证实了C12ORF40/REDIC1是一个新的重组蛋白,并综合运用多种方法揭示了它在CO形成中的关键作用,加深了对减数分裂重组调控的认识。然而,生理条件下REDIC1如何被募集到染色体轴以及如何与ZMM蛋白一起稳定重组中间体还不是很明确。后续研究工作还在继续推进中。
上一篇:
解析影响CD4 T细胞活化的因素
下一篇:
发现疟蚊肠道共生菌利用胞外囊泡跨界递送抗疟效应分子靶向杀灭疟原虫的新机制
已经到最底了
技术支持:
库价化学
Copyright © 2024北京螽斯羽生物有限公司 备案号:
京ICP备2023018288号-1