大脑的一个基本目标是提取世界的潜在结构,并对其因果关系进行建模。随着线索-结果关系的变化,大脑必须能够灵活地更新这些模型。但仍不清楚在群体水平上学习关联是如何表现的,以及这些表现是如何作为学习的功能而变化的。编码习得关联的一个重要区域是海马体(hippocampus,HPC)。遗传、解剖学和功能数据表明,啮齿动物HPC的背侧和腹侧分支(dHPC和vHPC)在学习世界时起着不同的作用[1,2]。dHPC中的神经元反应编码有关世界的相对具体的信息,vHPC表征对更抽象的元素做出反应。因此,在学习过程中,dHPC的详细表征可能支持基于局部线索的联想记忆的形成,而广泛的vHPC表征可能在多个经验中概括知识和/或赋予联想发生的意义。
虽然dHPC和vHPC可以编码所探索环境的独特特征,但这些区域在编码联想记忆时是如何参与的,在很大程度上仍然是未知的。此外,当学习关系被操纵和更新时,神经反应是如何转变的也不清楚。在这项研究中,当小鼠学会将气味刺激与食欲或厌恶结果联系起来时,使用dCA1或vCA1群体活动的双光子体内成像来跟踪相同神经元在多天内的活动。这使研究者能够研究任务相关信息是如何通过背腹侧(DV)海马轴表示的,包括这些表示如何随着学习而演变,它们随时间的稳定性等。
该研究使用双光子钙成像在几天内跟踪了相同的dCA1和vCA1神经元,以确定气味结果学习阶段的反应是如何演变的。最初,气味在dCA1中引起了强烈的反应,而在vCA1中,气味反应主要是在学习和嵌入有关配对结果的信息后出现的。这两个区域的种群活动随着学习迅速重组,然后稳定下来,即使在灭绝或与不同的结果配对之后,也能将习得的气味表征储存数天。此外,当小鼠预期行为控制下的结果时,该研究在CA1上发现了稳定、强大的信号。这些结果显示了海马体如何编码、存储和更新习得的关联,并阐明了背侧和腹侧海马体的独特贡献。
一、CA1中气味特征的表征
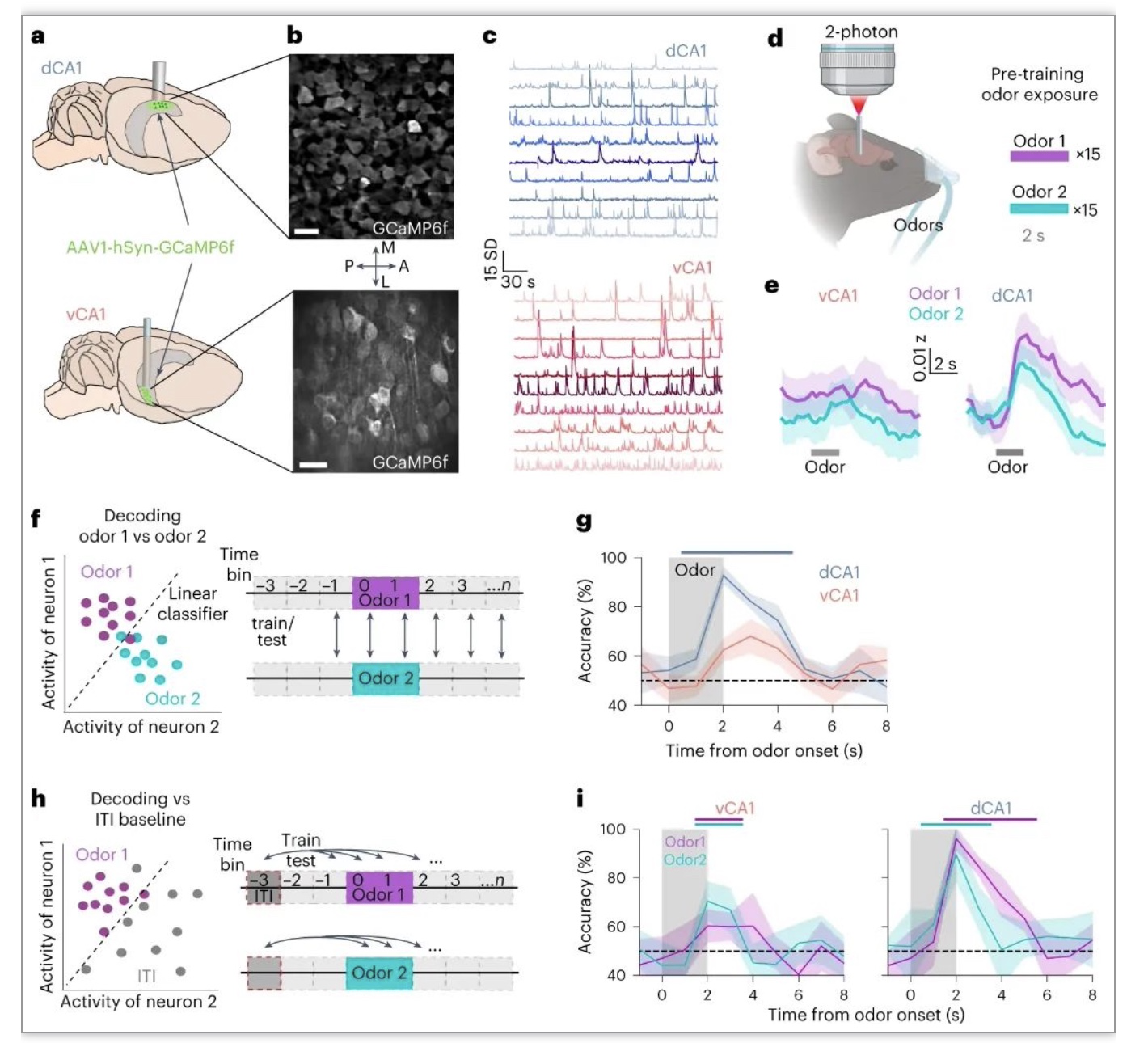
为了评估训练前对气味的基线反应,研究员首先记录了CA1对两种被动传递气味的反应。使用高分辨率双光子显微镜在表达钙指示剂GCaMP6f的小鼠中对dCA1和vCA1的神经反应进行了成像(图1a-c)。发现在dCA1中这两种气味都引发了强大的神经元反应(图1e)。为了检验种群对每种气味的反应是否不同,该研究训练了一个线性分类器来区分试验类型的活动模式(图1f)。解码分析表明,气味身份可以从dCA1群体活动中高精度解码(图1g)。气味诱发的群体反应也可以被准确区分(图1h,i)。相反,vCA1神经元的气味反应弱于dCA1(图1e),线性解码器在读取试验标识或从基线活动中区分气味诱发的活动时的表现明显差于dCA1(图1e-i)。总的来说,在最初接触缺乏行为相关性或有意义关联的气味时,气味特征在dCA1中表现强烈,但在vCA1中表现较弱。
图 1 在条件作用前,气味刺激在dCA1中比vCA1中表现得更强烈。
(图源:Biane JS,et al., Nat Neurosci, 2023)
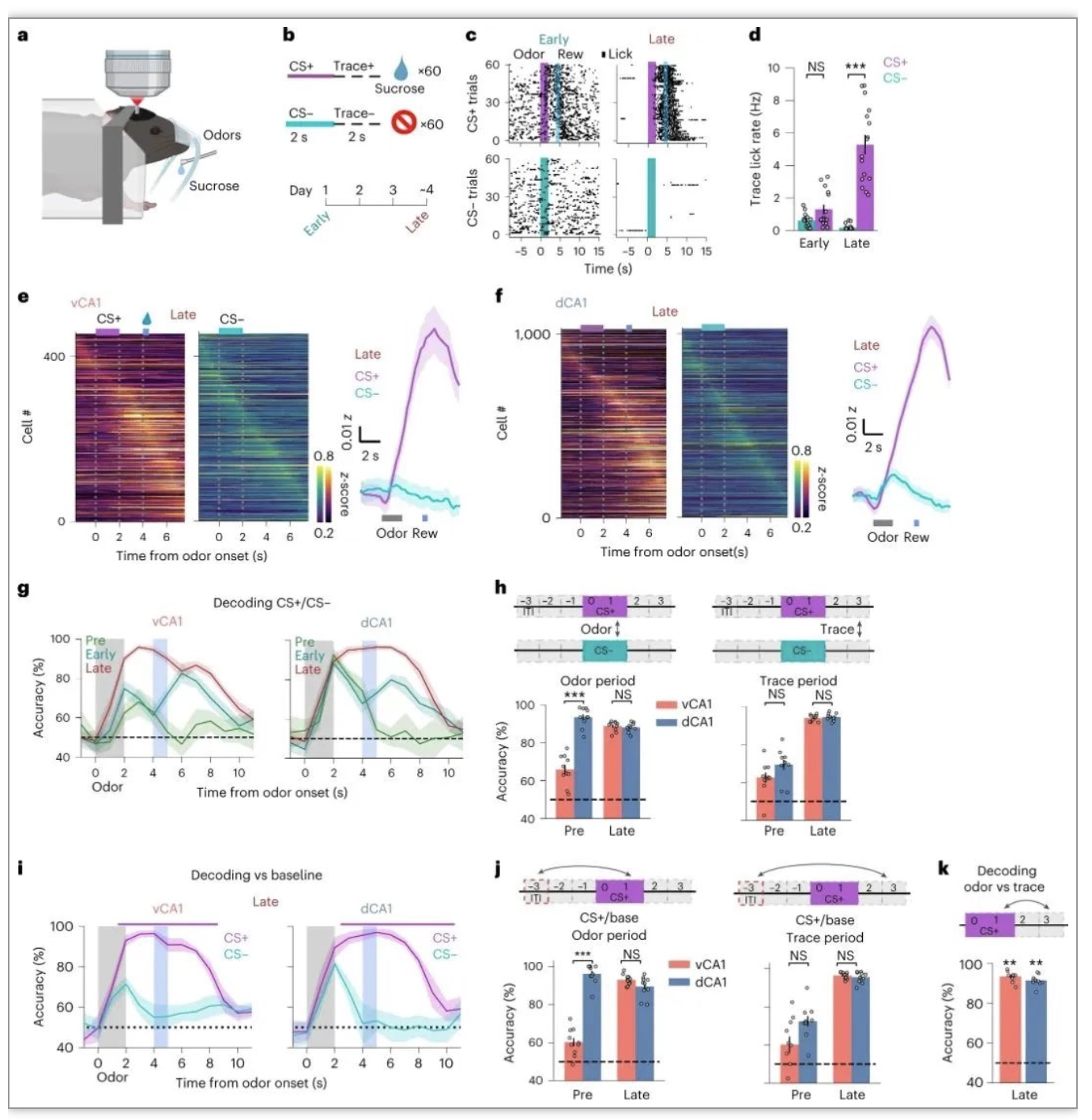
为了从潜在的奖励预期信号中消除气味表征的歧异,该研究使用了一种双气味痕迹食欲条件反射范式。经过大约4天的训练,在CS+追踪期间,小鼠表现出预期的舔舐,在所有其他任务期间,舔舐次数最少(图2a-d)。在vCA1中,气味-奖励关联的学习伴随着CS+气味呈现平均活动的整体增加(图2e),以及气味区分CS+与CS−种群活动的能力增强(图2g,h)。vCA1中试验型解码精度的提高似乎是由现在显著的CS+气味处理的改变所驱动的,因为气味反应(图2e)和解码气味周期与基线活动(图2i,j)都随着对CS+气味的学习而增加。因此,给刺激赋值会增加气味诱发的活动和腹侧CA1编码,这与dCA1相反(图2g,h)。随着学习,dCA1和vCA1的表征发生了平行变化(图2g-j)。dCA1和vCA1的平均示踪期诱发活性在CS+传递后显著增加,但在CS-传递后没有增加。总之,气味特征的表征存在于dCA1中,与学习无关,而vCA1中的表征则更依赖于习得的行为意义。
(图源:Biane JS,et al., Nat Neurosci, 2023)
在小鼠学会气味奖励关联后,从CS+试验中省略奖励。小鼠在第一次关联消失阶段的早期便迅速消除了条件反应(图3a-c)。第二天又恢复了条件反应, 动物迅速恢复了预期的舔舐行为,表明对奖励任务结构的记忆完好无损。相应的,vCA1的气味识别分类精度提高(图3d,e)。这些数据表明,vCA1中气味表征的辨别能力对与气味相关的当前值很敏感。
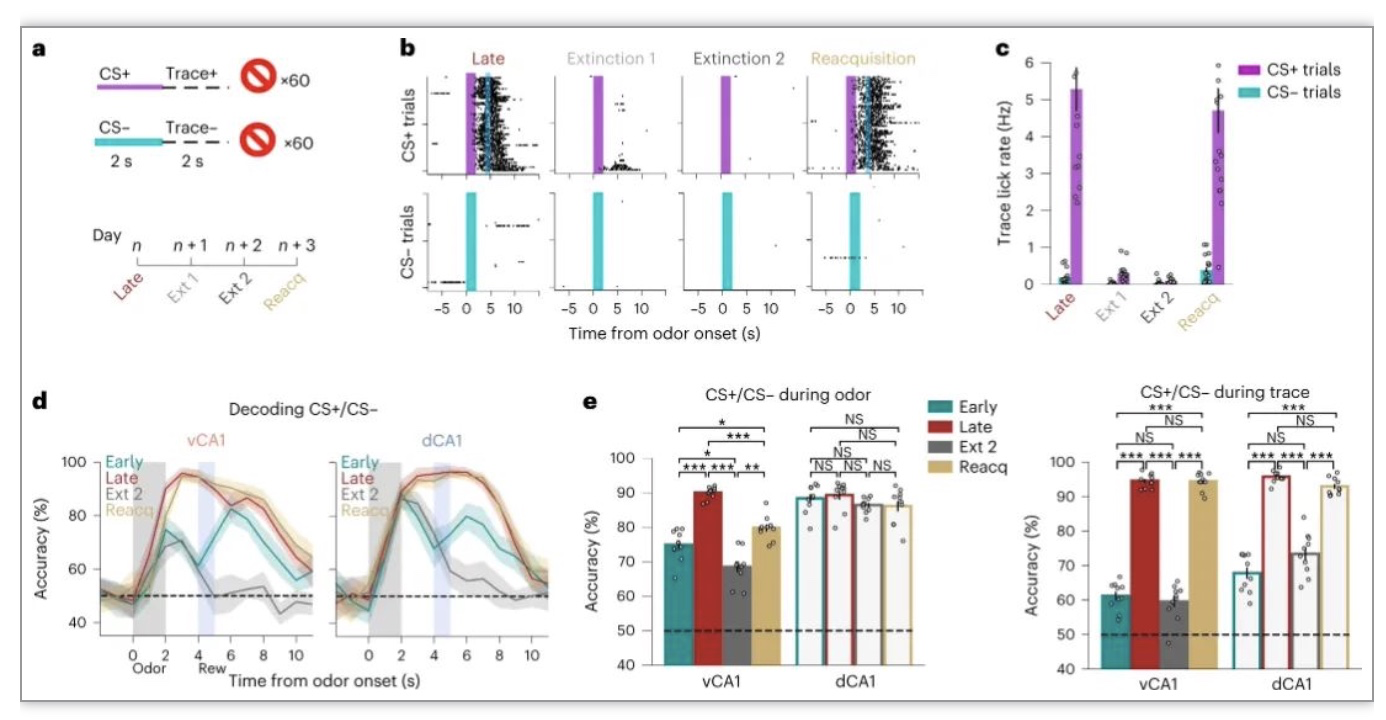
图3 习得的气味表征对消失很敏感,但可以迅速恢复。
(图源:Biane JS, et al., Nat Neurosci, 2023)
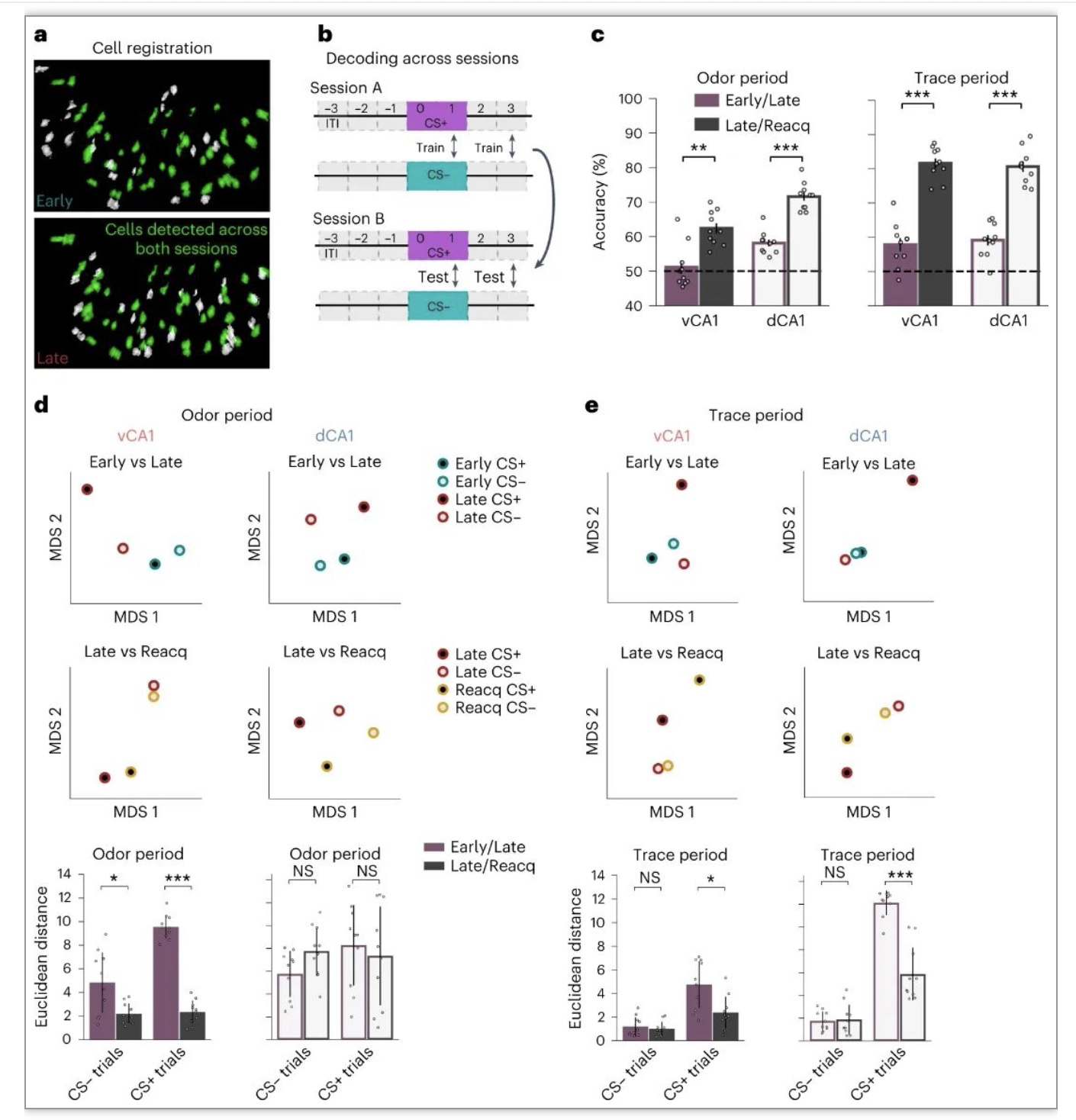
为了在训练的不同阶段探测气味和跟踪表征的稳定性,该研究对跨会话跟踪的神经元应用了跨会话分类器。结果表明,对于dCA1和vCA1来说,从早期到晚期会话使用跟踪周期活动的CS+/ CS−跨会话解码都很差,这表明随着学习,跟踪周期发生了变化(图4)。该研究进一步使用跨会话解码来检查单次学习到的任务表示的稳定性,发现vCA1和dCA1的气味周期和跟踪周期都保持相对稳定(图4c)。这与在初始学习期间观察到的任务表征的不稳定性形成对比,在几天内,在气味奖励偶然性的退化和恢复过程中都是稳定的。这些结果表明CA1可能是这些气味结果表征的存储位点。该研究还使用多维尺度(multidimensional scaling,MDS)可视化几何架构(图4d,e)。发现CS+表征和较小程度上的CS−表征随着初始学习(早期与晚期)而被修改,但它们在种群水平上仍然是动态的。
(图源:Biane JS,et al., Nat Neurosci, 2023)
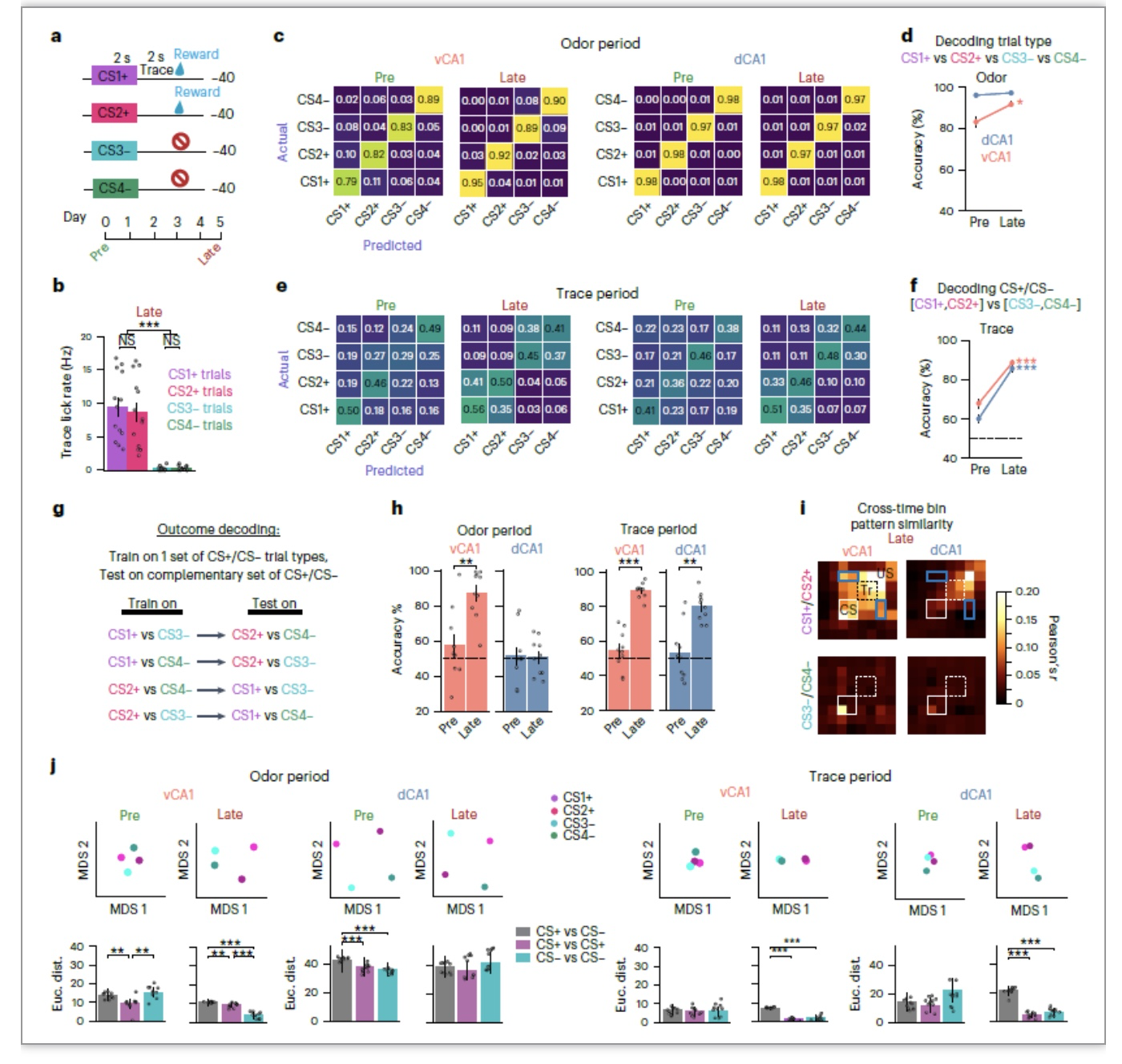
为了更好地解决“在学习期间,什么样的信息被编码”这个问题,该研究用四种气味刺激来训练小鼠:两只总是伴随着蔗糖奖励(CS1+和CS2+),两只没有任何结果(CS3−和CS4−)(图5a,b)。发现在学习后,dCA1和vCA1在气味传递期间的气味识别都可以被高精度地预测。在追踪期间,单个试验类型仍然是可区分的。因此,学习后,跟踪期活动在试验类别之间具有高度的可比性,但在每个类别内则不那么明显。在学习后, dCA1和vCA1在追踪期间的结果解码精度都很高(图5h),vCA1在气味期间的结果解码精度也很高,这表明,神经表征在dCA1中从编码线索身份转变为结果预期,而在vCA1中,预期结果在整个试验中表示,并在气味暴露期间与特定线索身份的信息进行多路复用。
图 5. 个体气味表征主导了dCA1,而vCA1包含关于未来结果的信息,两个区域都代表追踪期的预期结果。
(图源:Biane JS,et al., Nat Neurosci, 2023)
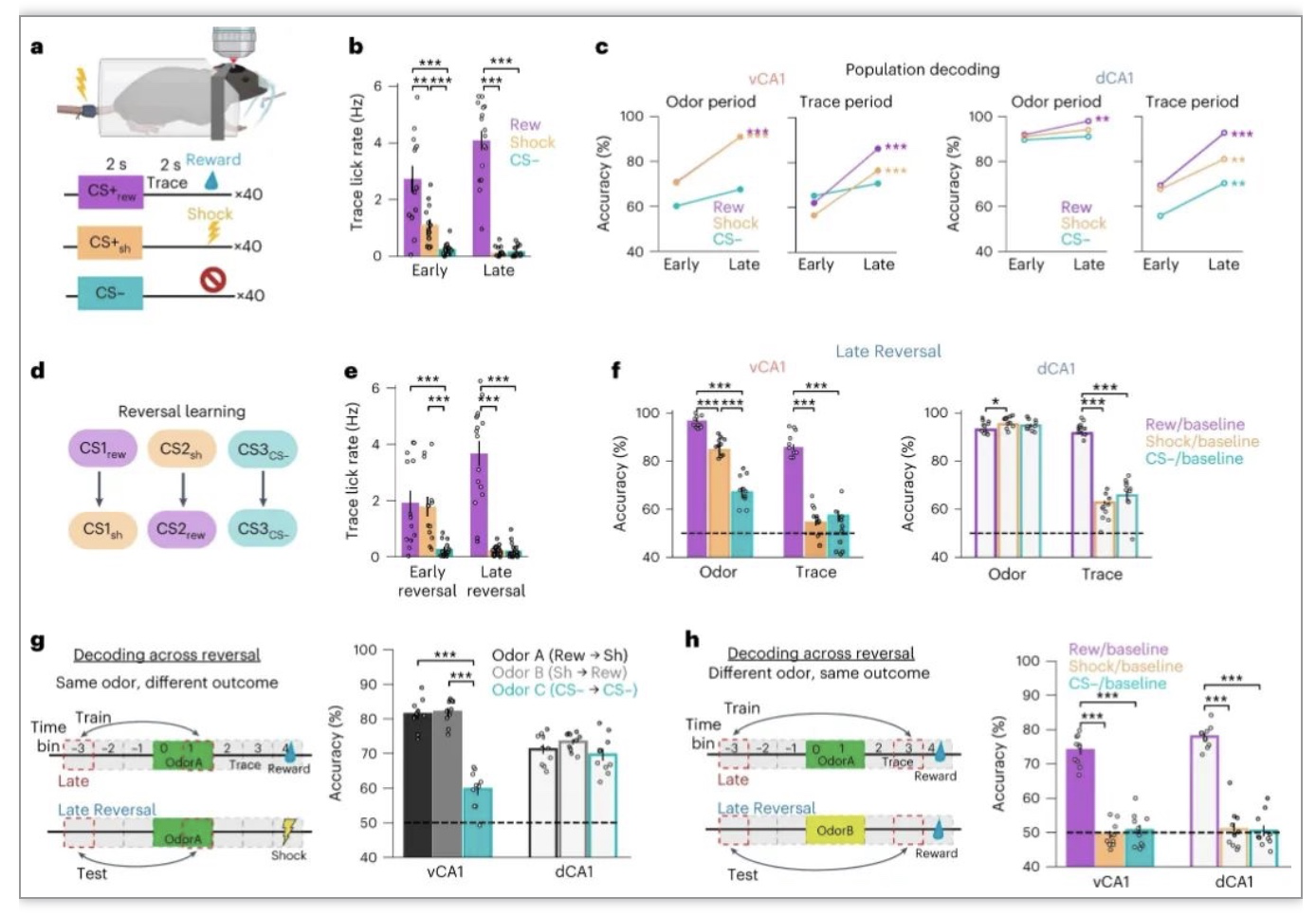
该研究使用三种新的CS气味训练小鼠进行联想学习任务,这些气味与蔗糖(CS+rew)、尾部电击(CS+sh)或无操作(CS−)配对。对气味期vCA1神经数据的分析显示,CS+sh与CS+rew的结果都显示随着学习气味编码的增加。然而,尽管在vCA1和dCA1的休克试验中,解码精度随着学习而提高,但与奖励试验相比,解码精度显著降低。随后研究者反转了偶发事件,即先前奖励的气味现在与电击配对,反之亦然。在vCA1和dCA1中,无论气味是否预示着蔗糖或休克,对特定气味的神经表征都保持完整。总之,这些结果表明,dCA1和vCA1中的气味表征与US的性质无关,并且在这些区域的学习中出现了奖励前的稳定信号(图6)。
(图源:Biane JS,et al., Nat Neurosci, 2023)
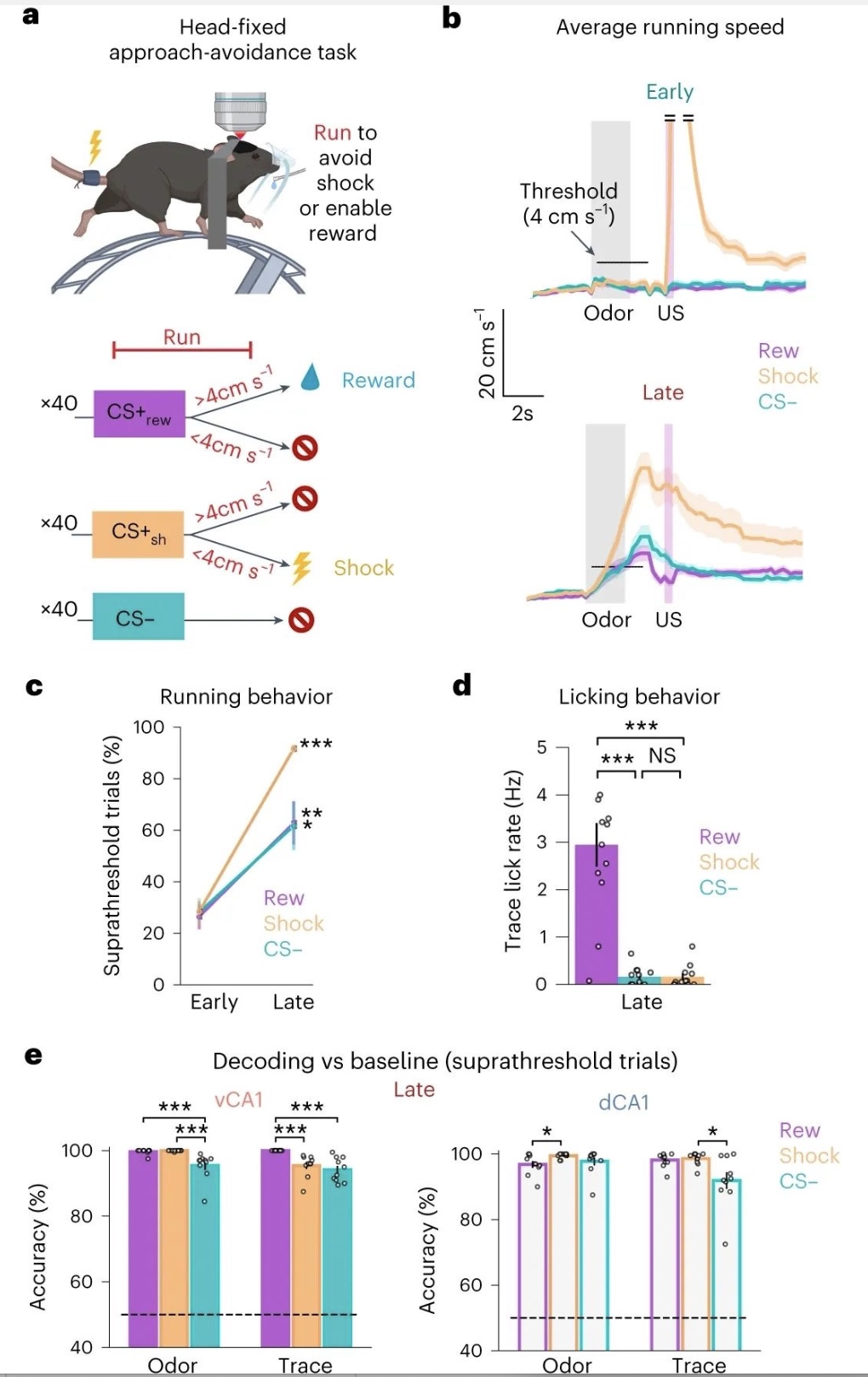
该研究推断dCA1和vCA1的奖赏前信号如此强烈的原因可能是这些结果在行为相关性上的差异:奖励试验需要操作性反应(舔),而电击传递是不可避免的。因此,该研究开发了一个头部固定的靠近-回避任务。在大约5天的时间内,小鼠学会了逃脱电击并能够进行奖励传递(图7)。
图7. 对结果的工具性控制增加了联想学习中的任务相关表征。
(图源:Biane JS,et al., Nat Neurosci, 2023)
这项研究展示了dCA1和vCA1相协调。HPC可以被认为是沿着DV轴进行从外部偏向到内部的环境变量编码的转变。这样的分工可以促进学习,并支持丰富的内部模型的创建[3,4]。此外,该简化模型与海马损伤研究一致,其中dHPC损伤影响陈述性记忆,vHPC功能障碍与正确分配密切相关,如创伤后应激障碍、成瘾和抑郁症[5,6]。该研究还发现 dCA1和vCA1中一些任务变量的表示方式有明显的重叠。由于dCA1和vCA1的传出连接模式差异很大,很可能每个区域都将此信息广播给不同的下游目标。当dCA1和vCA1的输出确实汇聚到同一区域时,这些输入可能会被不同的处理,就像奖赏学习中伏隔核的情况一样。未来研究的一个有趣问题是,dCA1和vCA1是否从公共或单独的输入源延续这些重叠,或者可能相互影响。最后,尽管之前的报道已经显示了各种“非运动”大脑区域的运动和神经活动之间的正相关关系,但dCA1和vCA1中神经活动与舔舐、呼吸或跑步行为之间的相关性大多较弱。未来的研究可以进一步阐明持续或任务诱发的行为,如姿势调整、奔跑以外的爪子运动或舔舐或呼吸以外的运动,如何影响海马活动。