18101298214
公司首页
实验手册
小分子化合物
细胞培养和检测
定制服务
文献资料
联系我们
订购指南
公司首页
实验手册
小分子化合物
细胞培养和检测
定制服务
文献资料
联系我们
订购指南
首页
实验手册
花青素苷在植物逆境响应中的作用
发布日期:2023/8/28 21:28:00
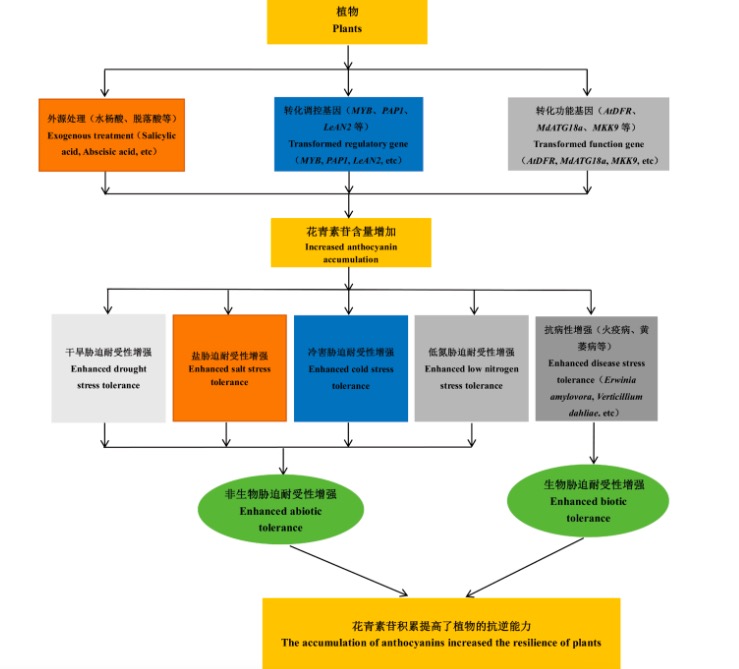
花青素属于类黄酮类化合物,是一类具有水溶性的天然色素。自然条件下,花青素以糖苷(花青素苷)形式存在。花青素苷的种类、含量和分布是植物花、茎、叶、果实等器官组织呈现不同色彩的重要原因。花青素苷在植物抵抗干旱、低温、盐胁迫、低氮等非生物胁迫及火疫病菌、软腐病菌、黄萎病菌等生物胁迫中具有重要的作用。
1 花青素苷的生物合成
1.1 花青素苷生物合成阶段
花青素苷的生物合成途径是黄酮类化合物合成途径中的一部分,属于次生代谢产物黄酮类的分支途径,主要受结构基因和调节基因的控制。黄酮类化合物在植物中主要呈现红色、蓝色和紫色。花青素苷生物合成途径的前体物质是苯丙氨酸,合成路径主要分为三个阶段。第一阶段是由前体物质苯丙氨酸经苯丙氨酸解氨酶 (PAL)、肉桂酸羟化酶 (C4H)、4-香豆酰辅酶A连接酶(4CL)等调控合成4-香豆酰辅酶A的过程;第二阶段,查尔酮合成酶( CHS)催化4-香豆酰辅酶A和丙二酰辅酶A合成黄色的查尔酮,再经查尔酮异构酶(CHI) 的异构化作用形成无色黄烷酮,再通过黄烷酮-3- 羟化酶(F3H)催化形成黄 烷酮和无色二氢黄酮醇;第三阶段,各种花青素苷 的生物合成阶段。黄烷酮和无色的二氢黄酮醇被二氢黄酮醇 4-还原酶(DFR)还原成无色的花青素苷。无色花青素苷在花青素合成酶(ANS)的催化作用下通过加氧反应生成有色花青素苷,然后通过类黄酮3-葡糖基转移酶(3GT/UFGT)的催化作用形成不同颜色的花色素苷。
1.2 结构基因
结构基因是花青素苷生物合成的功能性基因, 在植物花青素苷的合成中具有重要的作用,主要负责编码植物花青素苷生物合成途径中的各种酶,由上游基因群和下游基因群组成。上游基因群包括 CHS、CHI和F3H,主要负责编码合成通路起始位点上的参与黄酮、黄酮醇和糅红合成的相关酶类,这些结构基因的表达水平受花青素苷生物合成过程中的调节基因的激活或抑制进而决定花青素苷所积累的部位与水平;下游基因群包括类黄酮 3'-羟化酶基因 (F3'H)、DFR、 ANS、UFGT等,主要负责编码合成花青素苷和原花青素苷的相关酶类。
1.3 调节基因
调节基因是花青素苷生物合成的调控性基因, 在植物花青素苷的合成中具有重要的作用,主要包括三类转录因子,即MYB转录因子、bHLH转录因子和 WD40转录因子,对其合成途径中结构基因的表达具有重要的调控作用。这三类转录因子之间可以形成蛋白复合体,通过与植物花青素苷生物合成途径中结构基因启动子的顺式作用元件的结合来调控相关基因的表达。MYB 转录因子是植 物中最大的转录因子之一,在植物应对非生物胁迫 和生物胁迫中发挥着重要作用。bHLH 转录因子是植物体内的第二大类转录因子,在植物的生长发育、生理代谢及逆境应答过程中起着重要的作用,主要通过参与复杂的信号通路,改变大量下游基因的表达水平来实现对非生物胁迫的适应性应答。WD40 转录因子是一类古老的蛋白家族,其结构域是真核生 物中最丰富和高度保守的结构域之一,参与了许多植物细胞过程,如转录调控、细胞分裂、囊泡形成与转运、加工、信号转导等。
2 植物花青素苷在非生物胁迫响应中的作用
2.1 花青素苷在干旱胁迫响应中的作用
干旱胁迫作为最主要的非生物胁迫对植物的生长和发育具有很大影响,很多研究表明在干旱胁迫下,花青素苷合成相关的 结构基因和调节基因的表达水平上调,花青素苷的含量上升,植物的抗干旱能力显著提高。在转小麦 R2R3-MYB 型基因 TaPL1的拟南芥中,参与早期和晚期花青素苷生物合成步骤的结构基因上调表达并且花青素苷在叶片中的含量增加,从而提高转基因拟南芥的抗旱能力 。在过表达番茄SlyMYB7-Like基因的拟南芥中,花青素苷合成途径中关键酶基因的表达水平升高,导致花青素苷含量的提高,从而导致转基因拟南芥叶片颜色发紫。在干旱胁迫下,脱落酸处理草莓果实后,PAL,C4H和DFR的活性增强,花青素苷生物合成相关的转录因子 FaMYB1,Fab-HLH3 和草莓透明种皮 1 (FaTTG1)的表达水平上调,花青素苷的含量显著上升,推测花青素苷含量的增加 导致脱落酸处理后草莓果实对干旱胁迫的抵抗能力提高。在干旱胁迫下,拟南芥植株体内具有自由基清除活性的花青素苷的过度积累,导致其体内活性氧的积累量减少,从而增强了其对干旱胁迫的抵抗能力。综上所述,植物可以通过提高与花青素苷合成相关基因的表达水平促进花青素苷含量的积累,进而提高植物的抗旱能力。
2.2 花青素苷在盐胁迫响应中的作用
土壤盐胁迫对植物细胞膜透性、光合作用和呼吸作用等会产生重大影响,是植物生长发育及产量 形成的重要限制性因素之一。在转Del基因的烟草中,与花青素苷合成相关的基因NtCHS、NtCHI、 NtF3H、NtDFR和NtANS的表达水平显著上调,叶和花中的花青素苷含量显著增加,从而使转Del基因的烟草抗盐能力显著提高。在过表达 AtDFR 的甘蓝型油菜中,DFR基因的转录水平显著升高,花青素苷含量显著提高,从而使转基因甘蓝型油菜对盐胁迫的抵抗能力明显增强。尿苷二磷酸葡萄糖醛基转移酶UGT79B2 和 UGT79B3,可以将 UDP-糖基转移到 矢车菊素使其更稳定,拟南芥中过表达 UGT79B2/ B3 基因可以增加花青素苷的积累,提高转基因植物对盐胁迫的抵抗能力;另一方面,采用 RNAi 和 CRISPR - Cas9 基因编辑技术两种方法获得 UGT79B2 和 UGT79B3 基因的突变体,其花青素苷含量显著降低,抗盐能力显著低于野生型。过表达甘薯IbMYB1 基因的马铃薯中与花青素苷合成相关的基因(CHS, DFR, ANS)表达上调,花青素苷含量升高,从而使转基因马铃薯对盐胁迫的抵抗能力增强。转甜樱桃R2R3-MYB转录因子PacMYBA 的拟南芥中花青素苷含量升高,转基因拟南芥对盐胁迫的耐受性提高,推测转基因拟南芥的花青素苷 积累增加可能导致对盐胁迫的抵抗能力增强。因此,向植物中转入与花青素苷生物合成相关的结构基因、调控基因以及外源处理等,可以显著提高花青素苷合成相关基因的表达水平,增加花青素苷的积累,进而提高转 基因植物对盐胁迫的抵抗能力。
2.3 花青素苷在冷胁迫响应中的作用
低温冷害是影响我国农业生产的主要灾害之 一。农作物在其生长发育期间,由于遭受低于其生长发育所需的环境温度,会引起农作物生育期延迟,或使其生殖器官的生理机能受到损害,导致农业减产。过表达番茄LeAN2基因的转基因烟草,与野生型烟草相比,几种花青素苷生物合成相关基因的表达水平上升,花青素苷含量显著提高,进而增强了转基因烟草对冷害胁迫的抗性。过表达拟南芥UGT79B2 / B3基因显著提高了植物对低温的耐受性,UGT79B2 和 UGT79B3 的表达由 CBF1直接控制以响应低温,结果表明 UGT79B2和UGT79B3为编码花青素鼠李糖基转移酶基因,受 CBF1 调节,通过调节花青素苷积累来 提高植物对低温的耐受性。苹果中过量表达 MdbHLH33 基因可以产生红肉的愈伤组织,然后在愈伤组织中过表达Md⁃MYB15L基因可以降低花青素苷的合成水平,从而降低了其抵抗低温的能力;LBSMdMYB15L 基因是 MdMYB15L基因的结合位点序列,在过量表达Mdb⁃ HLH33 基因的红肉愈伤组织中过量表达 LBSMd⁃ MYB15L 基因,同样可以降低其抵抗低温的能力。因此,MdMYB15L基因在花青素苷诱导的抗低温胁迫中起到重要作用。因此,向植物中转入与花青素苷生物合成相关的结构基因或者调控基因,可以显著提高花青素苷合成相关基因的表达水平,增加花青素苷的积累,进而提高转基因植物对 冷害胁迫的抵抗能力。
2.4 花青素苷在低氮胁迫响应中的作用
氮是植物生长发育过程中必不可少的营养元 素之一,低氮会严重影响植物的生长发育,如叶绿素的发育、光合作用效率等。在低氮胁迫下,过表达 MdATG18a 基因的苹果中黄酮类物质合成基因和调控基因 MdCHI、MdCHS、MdANS、MdPAL、MdUF⁃ GT 和 MdMYB1 的表达量升高,转基因苹果植株的花青素苷的积累量增多,增强了其抵抗低氮胁迫的能力。拟南芥testa7-1 (tt7)突变体 是F3'H 基因缺失的突变体,在低氮胁迫下野生型为红色,突变体表现为绿色。拟南芥 testa7-1 (tt7) 突变体中过表达苹果MdF3'H基因,转基因植株花青素苷含量上升,植株颜色变红,表明花青素苷在拟南芥tt7突变体抵抗低氮胁迫的过程中具有重要作用。同时,在低氮胁迫下,拟南芥tt3突变体(DFR缺失)花青素苷合成关键酶DFR 合成受阻,存活率显著低于野生型,tt3突变体抗低氮能力减弱,结果表明花青素苷在拟南芥抵抗低氮胁迫的过程中具有重要作用。在低氮胁迫下,促分裂原活化蛋白激酶激酶 9( MKK9)基因的拟南芥,通过调控拟南芥的氮素获取和增加花青素苷积累,提高拟南芥对低氮胁迫的抗性。在拟南芥中,DELLA 蛋白通过与 PAP1 的 直接相互作用正向调节缺氮诱导的花青素苷积累, 增强其对花青素苷生物合成基因F3'H和DFR表达 的转录活性,提高拟南芥对低氮的抗性。以上结果表明植物可以通过调节花青素苷的含量来增强植物对低氮胁迫的适应性。
3 花青素苷在植物生物胁迫响应中的作用
生物胁迫包括病害、虫害和杂草等,这些生物胁迫严重影响植物的正常生长发育,近年来花青素苷在提高植物对生物胁迫的抗性方面已有很多研究。过表达玉米叶色基因的转基因苹果中,花青素苷的含量明显提高,并且对苹果黑星病菌和火疫病菌的抗性增强。转红色植物基因导致了棉花光诱导的花青素苷积累和棉花中的红/紫色色素沉着,从而增强了转基因棉花对棉铃虫和蜘蛛螨的抗性。从海岛棉中分离出病原体诱导的基因,该基因编码具有双加氧酶结构的花青素苷合成酶蛋白GbANS,GbANS优先在有色组织中表达,促进花青素苷的生物合成,增强棉花对黄萎病菌的抗性。拟南芥突变体 vsad1(即tt4)编码黄酮类化合物生物合成的关键酶 CHS受阻,vsad1突变体中黄萎病菌的浓度增加,并且在棉花植物中同源基因GhCHS的敲减增加了棉花对黄萎病菌感染的易感性,表明花青素苷可以提高拟南芥和棉花对黄萎病的抗性。用 疱疫病处理感疱疫病和抗疱疫病茶树的过程中,花青素苷生物合成相关基因发生差异性表达,花青素苷的含量发生变化,推测花青素苷在茶树品种抗细菌、真菌的过程中起重要作用。黑莓中的总酚、类黄酮和花青素苷可以延迟黑莓采后真菌的生长,提高了黑莓的抗真菌能力。以上结果表明花青素苷能够提高植物对生物胁迫的抗性。
上一篇:
动态RNA甲基化修饰及其调控机制研究进展
下一篇:
植物生长调节剂诱导无籽果实形成的作用机制
已经到最底了
技术支持:
库价化学
Copyright © 2024北京螽斯羽生物有限公司 备案号:
京ICP备2023018288号-1