果实的发育是一个由内源植物激素和外部环境因子共同调控的过程。无籽是果实一个重要经济性状,也是品种选育一项重要指标。未经受精作用而子房发育的单性结实和受精后种子败育都可以得到无籽果实。天然的无籽果实是在没有授粉或受精的情况下,子房中植物激素浓度达到促进果实发育阈值的结果。外源植物生长调节剂可以代替授粉和受精释放的信号,调控坐果和早期果实发育。
生长素是首个被发现能诱导单性结实的植物激素,在协调花到果实的转变(坐果)过程中起着关键作用。花前生长素响应因子(ARFs)与生长素/吲哚乙酸蛋白( Aux/IAA)形成的异二聚体复合物抑制生长素响应基因的转录,进而抑制子房生长。受精后子房中高水平的生长素激活泛素-蛋白酶途径(UPP)使Aux/IAA 降解,ARFs 被释放得以促进生长素响应基因的表达,启动果实发育。生长素作为最活跃的激素信号参与花到果实的转变,其含量在受精完成后急剧上升。转录组分析授粉后膨大的子房室细胞,发现参与合成、转运和响应生长素的基因在该组织中先表达,随后才启动赤霉素相关基因的表达。
2,4-二氯苯氧乙酸(2,4-D),萘乙酸( NAA)和吲哚乙酸( IAA)是三种常用于诱导无籽果实的生长素,其中2,4-D效果最显著。近年来在番茄和拟南芥上的研究中发现,在坐果的早期,外源植物生长调节剂使子房中产生了类似于授粉后的生长素积累,外源生长素、细胞分裂素处理后对生长素合成相关基因 IAA2、IAA8、IAA9、IAA14、IAA7和响应相关基因ARF8、ARF9、ARF4的表达均显著上调,却几乎不受外源赤霉素(GA3)的调控。
极性运输在生长素信号转导中起着至关重要的作用,在启动坐果和早期果实发育中也发挥着重要作用。生长素极性输出载体PIN-FORMED蛋白在质膜上的极性分布决定了生长素在细胞间的运输方向。通过RNAi技术沉默番茄的生长素外排蛋白4基因(PIN4)得到两个兼性单性结实的纯合株系L-2 和L-21。生长素极性运输抑制剂(NPA)处理番茄果柄,阻断生长素向下运输,使子房中IAA含量升高,从而诱导未授粉子房单性结实形成无籽果实。摘除茎尖也能诱导豌豆单性结实 ,而在切面上放置含有IAA或脱落酸的琼脂块则会扭转这种效应。这表明生长素在坐果过程中可能扮演着相反的作用,在子房中积累促进坐果,从茎尖极性运输抑制坐果。然而生长素的积累究竟是生长素合成增加、生长素降解减少或生长素转运改变的结果尚不清楚。
2 赤霉素
迄今为止被鉴定的136种赤霉素中,GA1,GA3,GA4,GA7和GA20已被用于诱导无籽果实,不同植物中各赤霉素诱导产生无籽的能力存在差异。在番茄中 GA3>GA1=GA4>GA20,但GA3处理的黄瓜和梨无籽率低,成熟前几乎全部脱落,效果远远不如 GA4+7。
赤霉素的生物学功能主要通过调控生物合成和信号转导来实现,任何一个过程或步骤的改变都将影响果实发育。赤霉素分解酶基因 (GA2ox1、GA2ox2、GA2ox3、GA2ox4 和GA2ox5)被沉默的转基因植株中,5%~37% 的未受精子房单性结实形成无籽果实。过表达赤霉素合成酶基因GA20ox1的转基因植株花期延迟,柱头突出,自花授粉受到阻碍,14%~30%未受精子房单性结实形成无籽果实。 上述转基因植株果实中,来自13-羟化途径的GA44、GA19、GA20和GA29的含量降低,而非13-羟化途径的 GA4及其活性前体GA9含量显著增加。在番茄中早期13-羟化途径产生的GA1是主要的活性赤霉素,GA4保持低水平。arf突变体中也表现出类似的现象,ARFs 表达下调可能激活非13-羟化途径,使 GA4 积累量明显上升。在果实发育过程中,13-羟化途径和非13-羟化途径可能只有一条起主要作用,当果实发育独立于受精作用时,另一条途径才起作用。
花前1~2周施加赤霉素,不去雄也能诱导产生无核葡萄。外源赤霉素诱导无核葡萄形成过程中,传粉3 d后可观察到花粉管尖端异常卷曲并停止生长,只有极少数能延伸至子房下部,没有花粉管能进入胚珠。在研究外源赤霉素诱导'巨峰'和'红地球'葡萄形成无籽果实机理时,在赤霉素处理25 d后观察到受精卵和胚乳核,随后受精卵和胚乳核出现异常并于30 d完全消失,这表明葡萄种子败育前花序已经授粉且成功受精。同时检测到过氧化氢和丙二醛的含量显著提高 ,超氧化物歧化酶、过氧化物酶、过氧化氢酶活性减弱 ,抗氧化酶基因 SOD1、SOD2、SOD3、CAT1、CAT2 和 POD 等表达下调。研究认为活性氧的水平与胚败育关系密切,SOD、POD、CAT 抗氧化酶活性下降和 MDA处于较高水平可能是外源生长调节剂诱导葡萄形成败育型果实的重要原因。
3 细胞分裂素
细胞分裂素在调控坐果和早期果实发育过程中发挥了重要作用。在分析番茄果实在不同发育阶段内 CKs含量时,发现在果实发育早期CKs的水平有两个峰值,第一个峰值出现在花期,tZR、iPR、DZ和DZR大量积累,不过这些CKs的积累与授粉无关,同时一些CKs生物合成的基因转录上调。花前高水平的CKs可能是未授粉子房生长或维持的必要条件,授粉后细胞分裂素分解相关基因 CKX1和CKX7的表达剧增而导致其含量快速下降。CKs的第二次峰值出现在开花后的第5天,细胞分裂素合成相关基因IPT1,2、CYP735A1, 2和LOG2的表达上调使tZ浓度达到最高,此时刚好是番茄果实细胞分裂的高峰期,这说明细胞分裂素tZ可能在授粉后果实发育中起着关键作用。
此外,外施细胞分裂素(CPPU)能诱导西瓜、瓠瓜、番茄、梨和黄瓜坐果并结出无籽果实,但其调控机制尚不清楚。比较单性结实与非单性结实番茄果实发现,细胞分裂素相关基因的转录水平在坐果期并没有显著差异。研究认为细胞分裂素诱导番茄单性结实可能部分依赖于增强赤霉素和生长素的生物合成。因此,细胞分裂素在诱导无籽果实中的作用还需进一步研究。
4 乙烯
乙烯在植物果实发育中的作用主要表现在成熟、衰老、脱落及对生物和非生物胁迫的反应。然而,目前的一些研究表明乙烯也会影响坐果和果实的早期发育。番茄子房转录组分析发现花期乙烯生物合成及信号转导相关基因大量表达,授粉或赤霉素处理诱导花向果实转变,使这些基因的表达在24h内减弱。研究发现在坐果和果实早期发育过程中生长素负调控乙烯的生物合成和信号,推测授粉后乙烯水平下降可能是生长素介导的结果。他们认为在雌花发育期间(开花前),乙烯控制雄蕊的停滞和花的适当成熟。也有研究认为乙烯可能与生长素和赤霉素拮抗调节子房休眠状态,但他们认为乙烯的作用是防止心皮在受精前发育成果实。
乙烯对果实定形和早期果实发育有负调控作用。乙烯在花中的减少不仅诱导雌花转变为两性花 ,而且促进果实的单性结实。乙烯不敏感突变或施加乙烯抑制剂(1-MCP)阻断乙烯信号,使番茄去雄的花直接发育成果实。四肽重复蛋白TPR1通过与CTR1竞争结合乙烯受体NR和ETR1,可以促进下游的乙烯信号响应,番茄中过表达TPR1可导致单性结实。此外,乙烯反应的增强会通过IAA9和SAUR1-like改变生长素反应。
果实的发育是多种植物激素协同调控的结果。对大多数植物而言,仅对未授粉子房施加单一激素,如生长素或赤霉素都不能得到正常果实,同时喷施这两种激素诱导产生的果实无论在外形还是组织形态上都和受精后得到的果实非常相似。生长素相关基因IAA9缺失的单突变体entire和赤霉素负调控因子DELLA缺失的单突变体procera产生无籽果实的同时,常伴随着畸形的胎座、发育不完全的子房室和假胚胎,但是两种激素相关的双突变体可有效改善果实品质与大小。
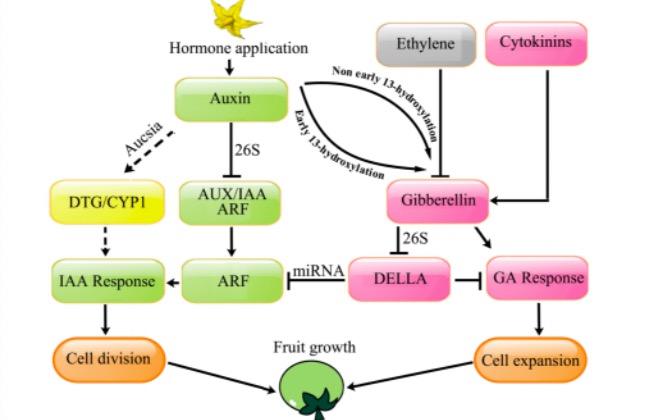
赤霉素是调控植物单性结实的关键激素,赤霉素可在不影响生长素信号相关基因(ASB1-like,
YUCCA2和YUCCA6)表达的情况下启动果实的发育。果实的发育取决于赤霉素的水平,外施植物生长调节剂导致子房中赤霉素的积累,进而启动坐果。细胞分裂素 CPPU 诱导番茄单性结实并提高了GA1+3和IAA的积累,上调GA合成酶基因 SlCPS 、SlGA20ox1、SlGA20ox2、和SlGA20ox3的表达,下调了 GA 分解酶基因的表达。多效唑在植物体内可以抑制GA的生物合成,外源施加PAC会显著抑制生长素和细胞分裂素诱导的单性结实,而这一抑制作用又可以被外源施加的GA3消除,赤霉素可能是生长素和细胞分裂素调控坐果的媒介。
解析内源激素启动果实发育的分子机制,是通过生物技术调控植物激素合成获得无籽株系的关键,也是进一步应用植物生长调节剂在更多作物种类诱导产生具有高市场价值无籽果实的科学基础。对拟南芥、葡萄和番茄无籽果实的研究表明,植物生长调节剂诱导产生无籽果实都有相似的途径。生长素信号通路的激活似乎是果实发育的第一步,生长素和细胞分裂素可能通过上调赤霉素合成基因并抑制分解和钝化基因,提高子房中活性赤霉素的含量从而调控果实发育。

