正常妊娠是提高出生率及出生人口素质的首要前提,但临床上复发性流产(Recurrent spontaneous abortion,RSA)的发生严重损害育龄妇女的生殖健康,并且影响出生率。人子宫内膜基质细胞(human endometrial stromal cells,hESCs)的蜕膜化对于妊娠建立和维持至关重要,蜕膜化过程伴随着基质细胞增殖、分化和凋亡,三者的平衡有利于维持恰当的蜕膜化,而蜕膜化稳态的异常会导致流产的发生[1],表现为蜕膜基质细胞(decidual stromal cells,DSCs)凋亡/过度焦亡[2-5]。JAZF1曾被报道与复发性流产相关,同时既往研究表明其表达水平下调会引起多种类型细胞凋亡,如胰岛β细胞[6]等。然而,JAZF1在蜕膜化稳态调控和复发性流产发生中的作用和机制尚未阐明。
该研究揭示了复发性流产蜕膜基质细胞中存在JAZF1表达下调、G0S2表达升高、细胞凋亡异常激活以及蜕膜化缺陷的情况;在体外细胞实验中,证实了JAZF1通过限制转录因子Purβ的活性从而抑制G0S2转录,参与人子宫内膜基质细胞的存活和蜕膜化的调节。这一发现在复发性流产的病理学和开发新治疗策略方面具有临床意义。
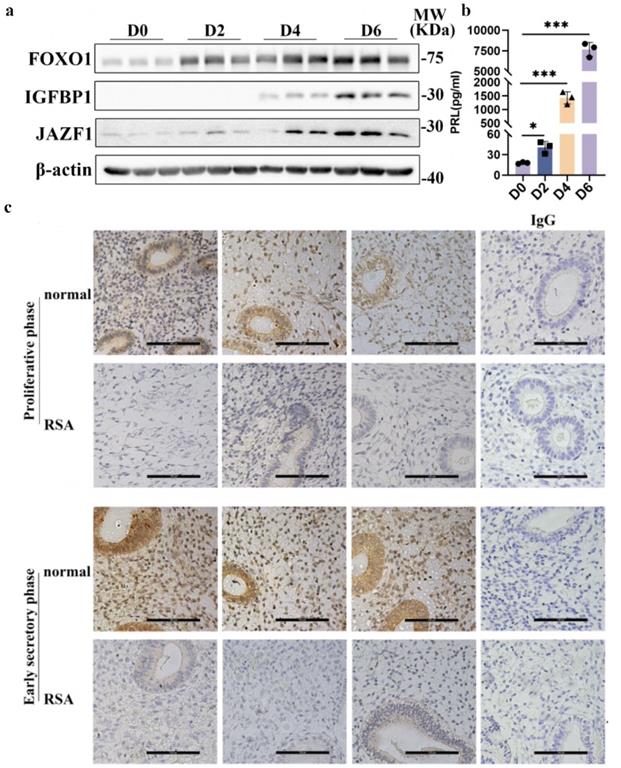
通过分析课题组前期获得的早孕蜕膜组织10×单细胞转录组测序数据集,作者发现JAZF1在RSA组的蜕膜基质细胞中均显著下调(图1a)。随后,作者检测了 RSA患者及正常妊娠妇女早孕期蜕膜组织以及原代蜕膜基质细胞中JAZF1的表达水平。结果提示JAZF1的水平在RSA蜕膜组织以及蜕膜基质细胞中均明显下调(图1b-d)。
为了确定JAZF1在蜕膜化过程中的表达,作者检测了蜕膜化前后永生化HESCs中JAZF1的表达水平,发现随着蜕膜化进展,JAZF1的表达均逐渐增加(图2a-b)。作者进一步检测了增殖期以及分泌早期的人子宫内膜组织中JAZF1的表达情况,提示JAZF1蛋白丰度在子宫内膜的分泌早期高于增殖期,尤其是在基质细胞中(图2c)。此外,结果也提示与对照组相比,JAZF1在RSA患者的增殖期以及分泌早期子宫内膜中表达水平均明显下调(图2c)。这些结果表明,JAZF1可能在蜕膜化的建立和发展以及RSA发生中发挥潜在调节作用。
图2 JAZF1在体外蜕膜化和月经周期中的动态表达模式
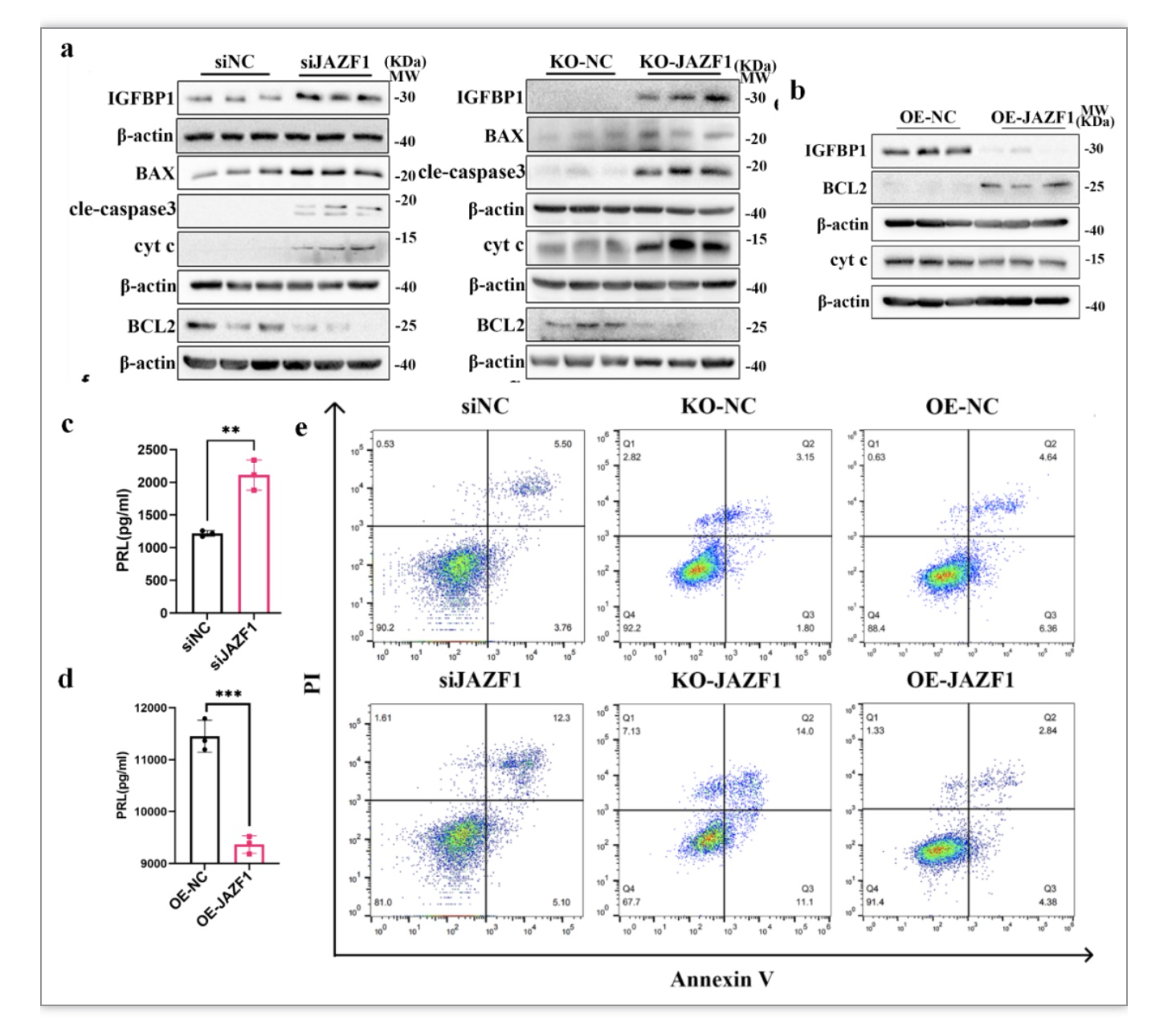
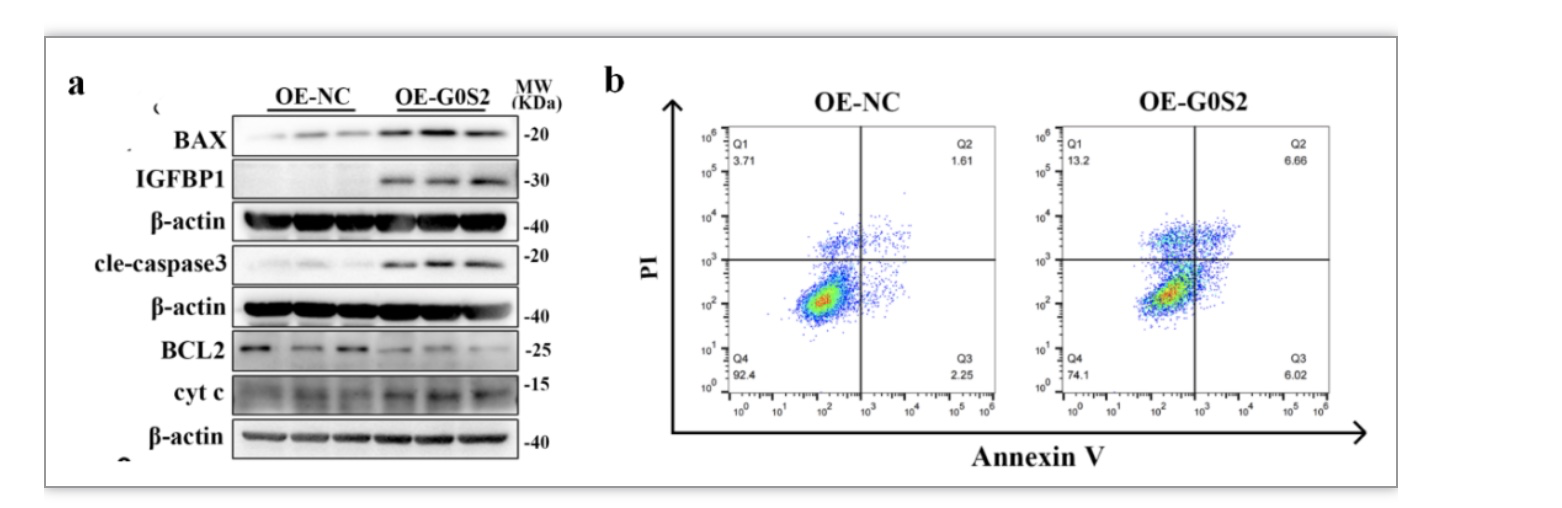
为了探究JAZF1在基质细胞中的功能,作者在HESCs中敲降/敲除JAZF1后进行体外蜕膜化,检测细胞凋亡以及蜕膜化相关指标表达情况。结果显示敲降或者敲除JAZF1后,促线粒体凋亡相关分子BAX、cyt c、cleaved-caspase3表达上调,而抗凋亡分子BCL2下调(图3a、c)。令人惊讶的是,JAZF1敲降或者敲除显著上调蜕膜化标志物PRL和IGFBP1的表达水平(图3a、c)。而过表达JAZF1能够下调IGFBP1、PRL以及cyt c的水平,以及上调BCL2(图3b、d)。流式细胞术也提示基质细胞中JAZF1缺失诱导细胞过度凋亡,而过表达JAZF1抑制细胞凋亡(图3e)。
图3 干预JAZF1后HESCs凋亡以及蜕膜化水平的情况
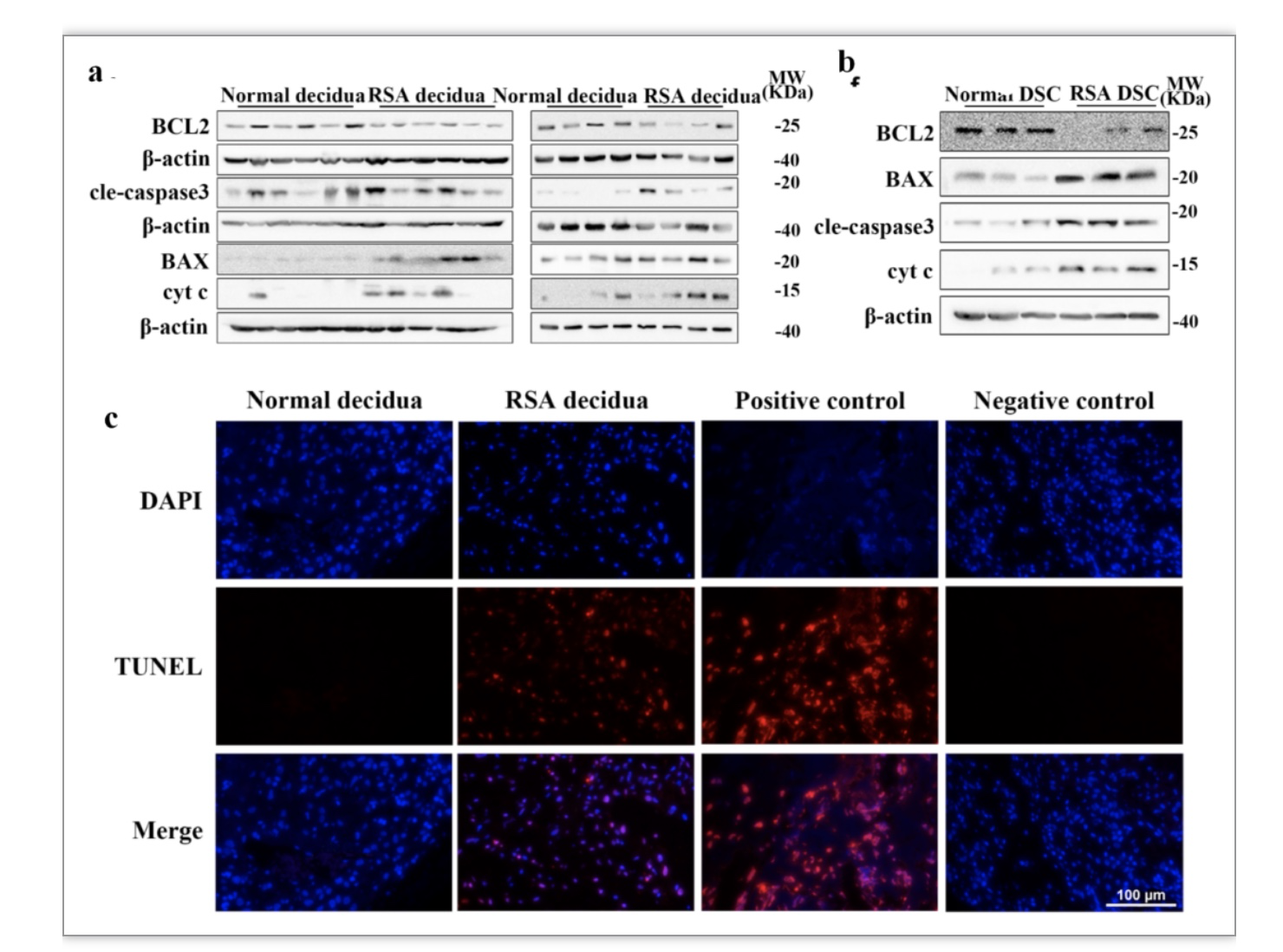
鉴于在体外蜕膜化过程中观察到的JAZF1下调诱导的促凋亡作用,作者进一步评估了RSA和对照组的蜕膜组织以及原代DSCs中的凋亡水平。结果显示RSA组蜕膜组织以及原代DSCs中促线粒体凋亡相关分子BAX、cyt c、cleaved-caspase 3明显上调,而抗凋亡基因BCL2水平下调(图4a-b)。同时,TUNEL染色法显示RSA组蜕膜组织的TUNEL荧光强度高于对照组,提示RSA蜕膜组织的凋亡水平较高(图4c)。
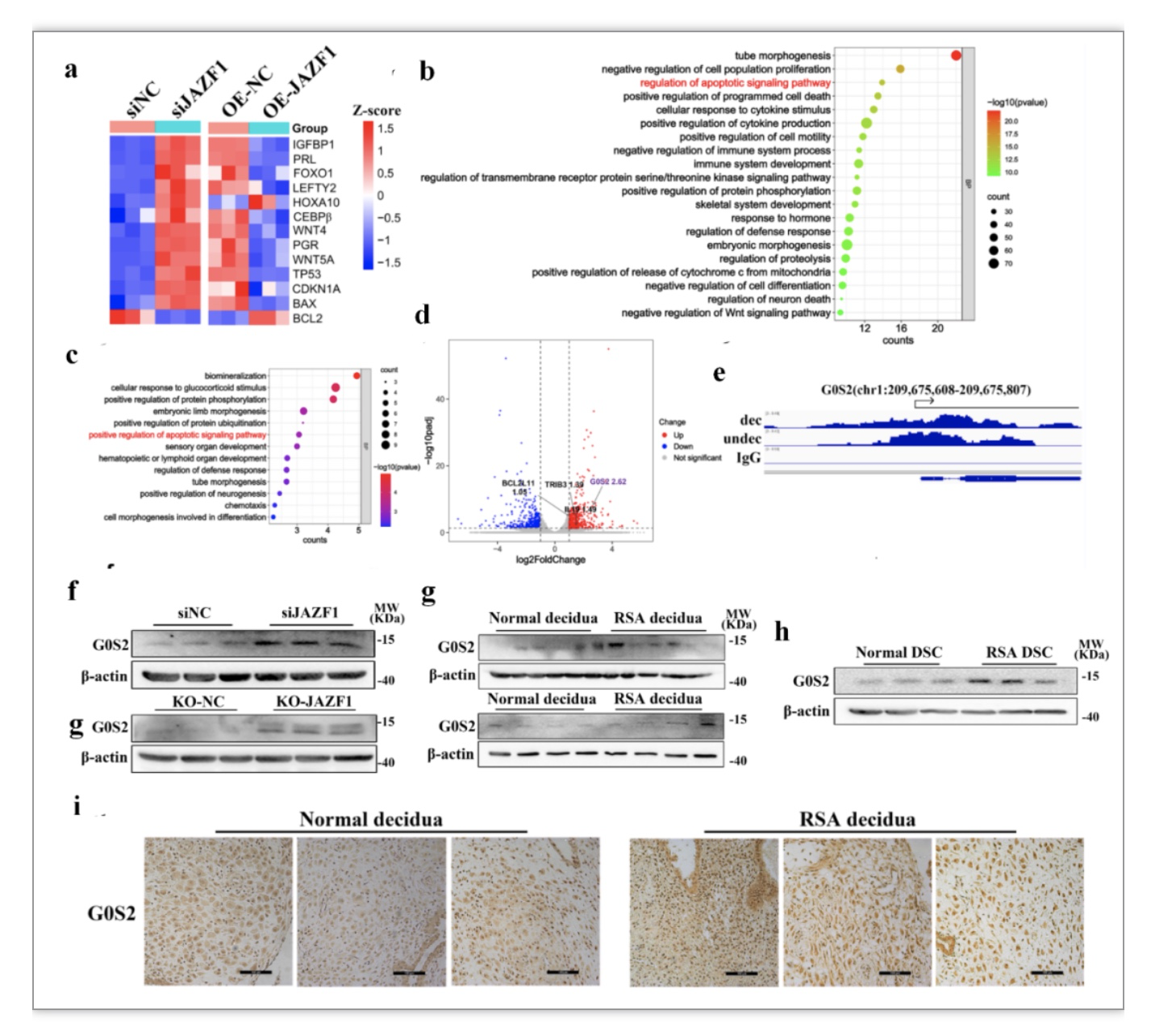
聚类分析分析结果显示敲降JAZF1后,促凋亡基因TP53、BAX、CDKN1A表达上调,抗凋亡基因BCL2下调;同时蜕膜化相关基因的表达显著上调,包括IGFBP1、PRL、FOXO1、LEFTY2、WNT4、PGR、WNT5A等(图5a)。GO功能富集分析提示,敲除JAZF1后上调的差异基因与凋亡信号通路的调节有关(图5b)。为进一步挖掘JAZF1在基质细胞中转录调控的关键靶点,作者将CUT&Tag和RNA-seq的数据进行了联合分析。GO分析提示差异基因仍富集在细胞凋亡激活的信号通路上(图5c),其中富集的基因G0S2在敲降JAZF1后的差异倍数最大,为2.52(图5d-e)。CUT & Tag-seq数据可视化图表明在G0S2的启动子区域存在潜在密集的JAZF1结合峰(图5e)。随后,在体外验证了JAZF1敲除后,G0S2水平显著上调(图5f)。同时,作者发现RSA蜕膜基质细胞中G0S2表达明显上调(图5g-i)。
图5 JAZF1抑制蜕膜化HESCs中G0S2的转录
既往研究表明,过表达G0S2诱导M1型巨噬细胞凋亡水平上调[7]。为了探究敲降JAZF1是否通过激活G0S2诱导hESCs过度细胞凋亡,作者在永生化HESCs中过表达G0S2并诱导蜕膜化,检测细胞凋亡以及蜕膜化水平。结果提示G0S2的过表达显著增加促线粒体凋亡相关分子BAX、BAX/BCL2、cleaved-caspase3和cyt c的表达,下调抗凋亡分子BCL2的表达(图6a)。流式细胞术也提示过表达G0S2会诱导细胞凋亡(图6b)。
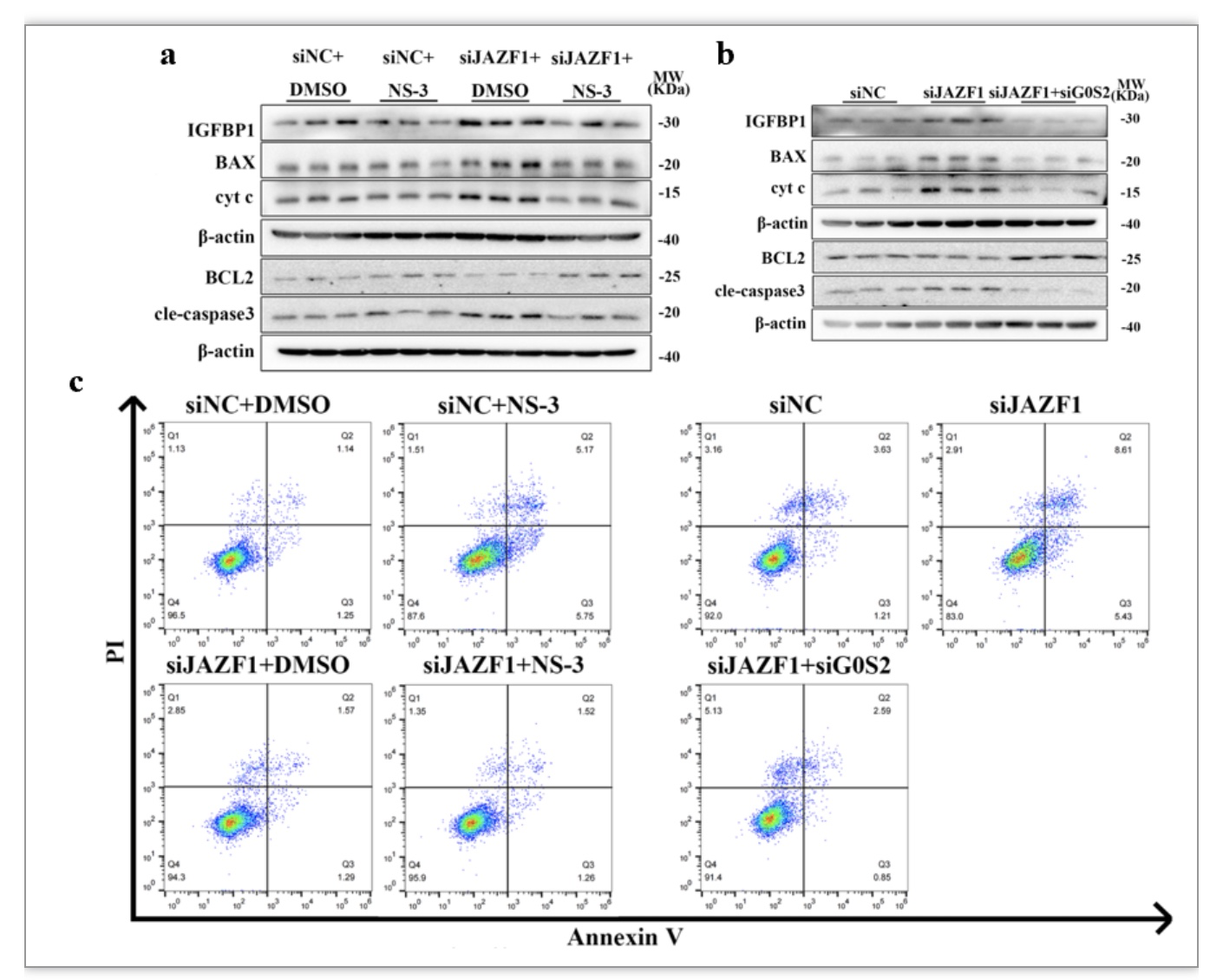
为了探索敲降G0S2能否改善由JAZF1缺失所引起的基质细胞过度凋亡和蜕膜化缺陷,作者在HESCs中敲降JAZF1同时利用siRNA或化学抑制剂NS-03-008抑制G0S2表达,并诱导蜕膜化6天后收集细胞检测细胞凋亡以及蜕膜化水平。结果显示蜕膜化过程中抑制或敲降G0S2下调了JAZF1敲降细胞中高水平的促线粒体凋亡相关分子BAX、cleaved-caspase 3和cyt c的表达,并上调了抗凋亡分子BCL2的表达(图7a-b)。此外,抑制或敲降G0S2下调了因敲降JAZF1所引起的IGFBP1的过度表达(图7a-b)。流式细胞术检测结果显示抑制G0S2能有效地挽救敲降JAZF1细胞的高凋亡水平。上述结果表明JAZF1的缺失通过激活G0S2诱导了蜕膜化hESCs的凋亡和IGFBP1表达缺陷。
图7 干预G0S2后对JAZF1敲降基质细胞的凋亡以及蜕膜化影响
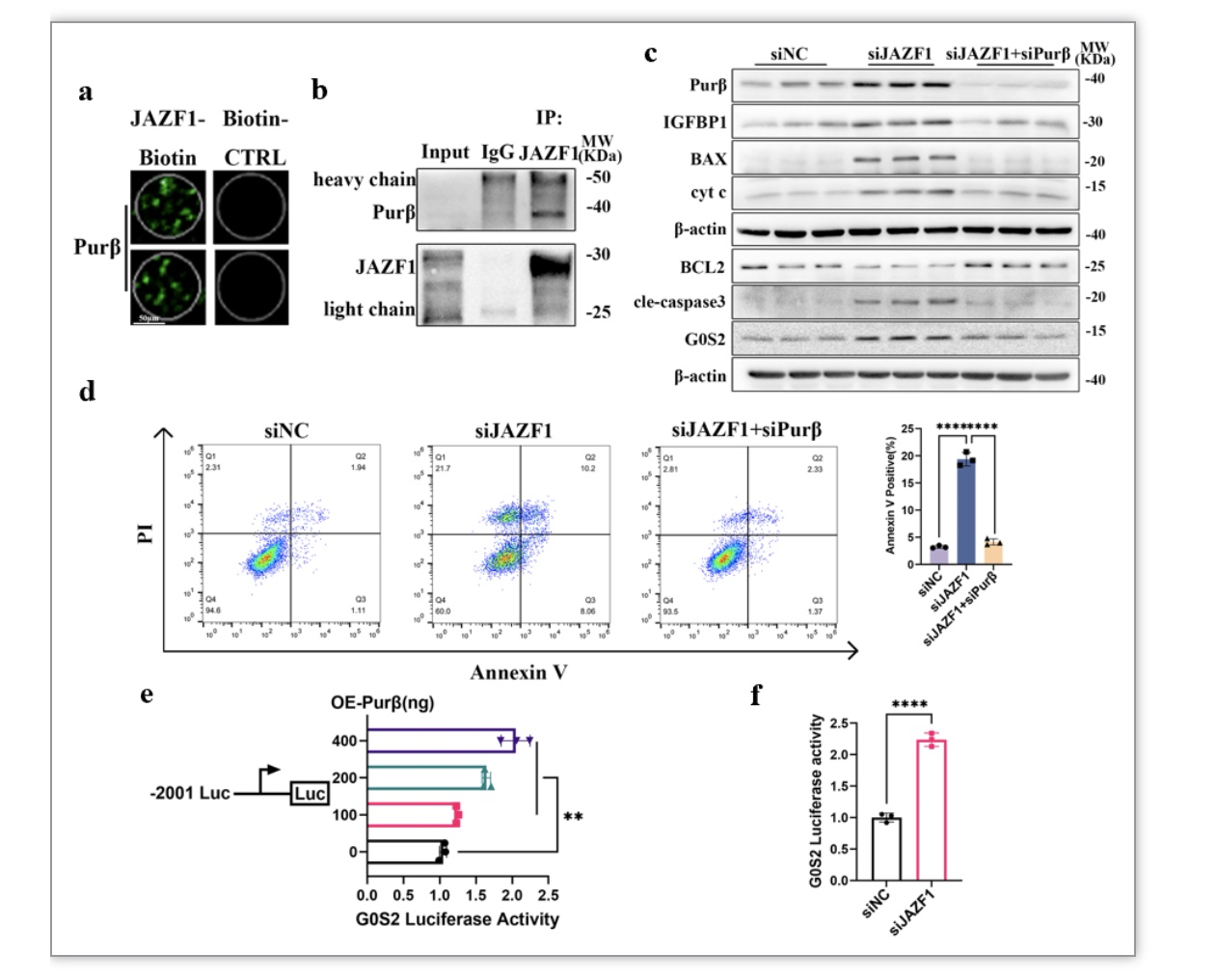
为了进一步明确JAZF1如何调控G0S2的转录水平,我们通过HuProtTM人类蛋白质组芯片、蜕膜化HESCs免疫沉淀以及质谱分析探索与JAZF1相互作用的蛋白。蛋白芯片结果提示JAZF1能与转录因子Purβ结合(图8a),同时MS结果和体外co-IP实验进一步证实了JAZF1和Purβ在蜕膜化HESCs中存在相互作用关系(图8b)。为了进一步确认Purβ在HESCs中的功能以及它是否参与调节G0S2的转录,作者在HESCs中共敲降JAZF1和Purβ后,诱导蜕膜化6天后收集细胞检测细胞凋亡以及蜕膜化水平。结果显示敲降Purβ后,有效地降低因JAZF1缺失导致的高水平的促线粒体凋亡相关分子BAX、BAX/BCL2、cleaved-caspase 3和cyt c的表达和高凋亡水平,并上调抗凋亡分子BCL2(图8c)。此外,敲降Purβ明显下调了敲降JAZF1引起的IGFBP1和 PRL的过度表达(图8c)。接下来,双荧光素酶报告基因结果显示,Purβ以剂量依赖的方式增加了G0S2启动子的荧光素酶活性(图8e)。敲降JAZF1后,Purβ显著上调了G0S2启动子的荧光素酶活性,这表明JAZF1抑制了Purβ对G0S2转录激活的功能(图8f)。
图8 JAZF1通过限制Purβ激活G0S2转录的功能来维持正常的蜕膜化
综上所述,本研究鉴定了JAZF1为蜕膜基质细胞凋亡的负调控因子,并发现其在复发性流产患者蜕膜基质细胞中的特异性下调。通过多种技术表明JAZF1的缺失可通过增强Purβ与G0S2启动子的结合,从而促进G0S2的转录,激活线粒体凋亡途径,引起蜕膜化标志物IGFBP1、PRL过度表达,诱发RSA患者人子宫内膜基质细胞过度凋亡和蜕膜化缺陷。本研究的发现为过度细胞凋亡与妊娠丢失之间的关系提供了重要的新知识,可能为维持蜕膜稳态和成功妊娠的新的调节途径,这一发现在复发性流产的病理学和开发新治疗策略方面具有临床意义。
本研究也存在几点不足。首先,由于RSA病因学的复杂性和异质性,本文的样本量有限。此外,蜕膜化涉及到蜕膜组织中的多种细胞类型,因此不能排除其他细胞对蜕膜化的贡献。最后,本研究的结果仅在细胞水平上得到了验证,未来使用条件基因敲除动物模型将有助于验证我们在JAZF1敲降基质细胞中的观察结果。