18101298214
公司首页
实验手册
小分子化合物
细胞培养和检测
定制服务
文献资料
联系我们
订购指南
公司首页
实验手册
小分子化合物
细胞培养和检测
定制服务
文献资料
联系我们
订购指南
首页
实验手册
研究表明CD8+T细胞暴露肿瘤抗原数小时内建立功能障碍印记
发布日期:2023/8/17 15:08:00
急性感染期间,
naive CD8
+
T细胞
经历激活并克隆扩增,进而能够产生效应细胞因子TNF-α、IFN-γ以及颗粒酶B和穿孔素[1]。慢性感染时,CD8
+
T细胞类似经历克隆扩增并发挥效应功能,但随着抗原持续刺激,CD8
+
T细胞增殖和效应功能受损,抑制性受体PD-1和LAG3等上调,呈现耗竭表型[2]。先前研究发现在肿瘤进展中,特异性CD8
+
T 细胞数天至数周内持续暴露于肿瘤抗原和免疫抑制肿瘤微环境,也表现出效应功能受损,抑制性受体上调[3-5]。然而,
肿瘤特异性CD8
+
T 细胞如何被激活、失去效应功能以获得功能障碍/衰竭的表观遗传特征尚不清楚,也未在体内进行评估。
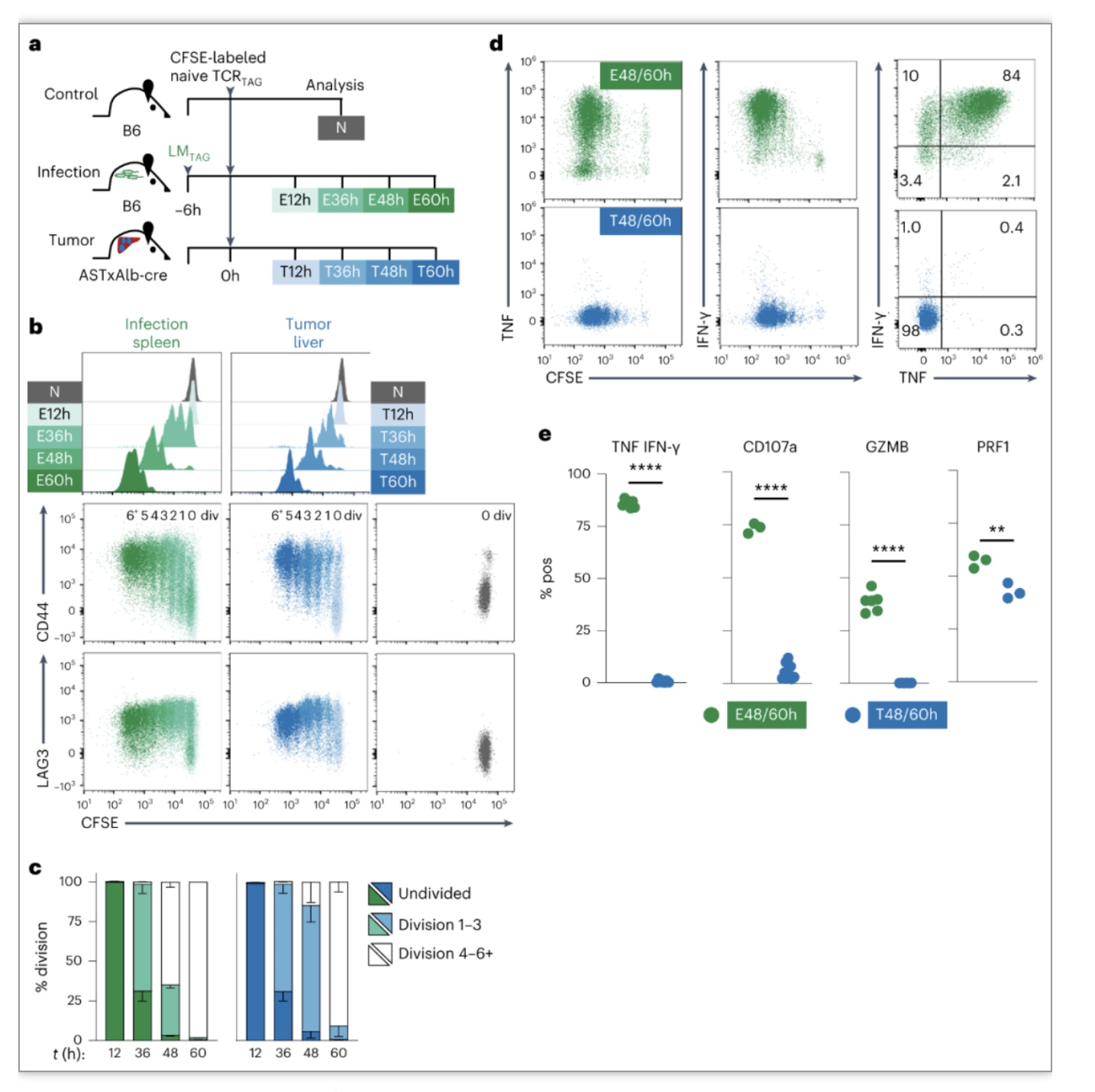
作者通过CFSE标记naive TCR
TAG
(Thy1.1)追踪CD8
+
T细胞增殖分化,并将它们分别回输到B6 (Thy1.2), LMTAG-infected B6 (Thy1.2) (表达TAG表位的LM
TAG
感染小鼠)和ASTxAlb-Cre (Thy1.2) (肝癌小鼠)小鼠体内。实验12、36、48和60小时重新分析受感染的脾脏或肿瘤肝脏中TCR
TAG
,发现荷瘤小鼠(T)和感染模型小鼠(E)中的T细胞都经历了非常剧烈的细胞分裂h和扩增。E-TCR
TAG
和T-TCR
TAG
均上调激活标记CD69和CD44以及TCR信号传导分子LAG3和PD-1。但是离体TAG肽再刺激后,T-TCR
TAG
无法产生效应因子IFN-γ和TNF,而受感染小鼠肝脏或脾脏E-TCR
TAG
能较强的产生效应细胞因子。以上数据表明
肿瘤抗原
促进
CD8
+
T细胞活化和增殖,但效应功能
受损
。
图1. CD8
+
T细胞接触肿瘤抗原后迅速活化和增殖,但效应功能受损
接下来作者进一步分析了肿瘤特异性CD8
+
T细胞效功能损伤发生的时间。通过分析CD69、CD44、LAG3和PD-1表达发现,与感染小鼠相似T-TCR
TAG
在6小时内被激活。在6小时E-TCR
TAG
开始产生效应细胞因子,但T-TCR
TAG
产生TNF能力受损,并且不产生IFN-γ,到12小时几乎完全无法产生两种细胞因子。表明
与感染激活的CD8
+
T细胞相比,
肿瘤特异性
CD8
+
T细胞效应功能在激活后数小时内受损
。
图2. 肿瘤特异性CD8
+
T细胞效应功能损伤发生在细胞分裂前
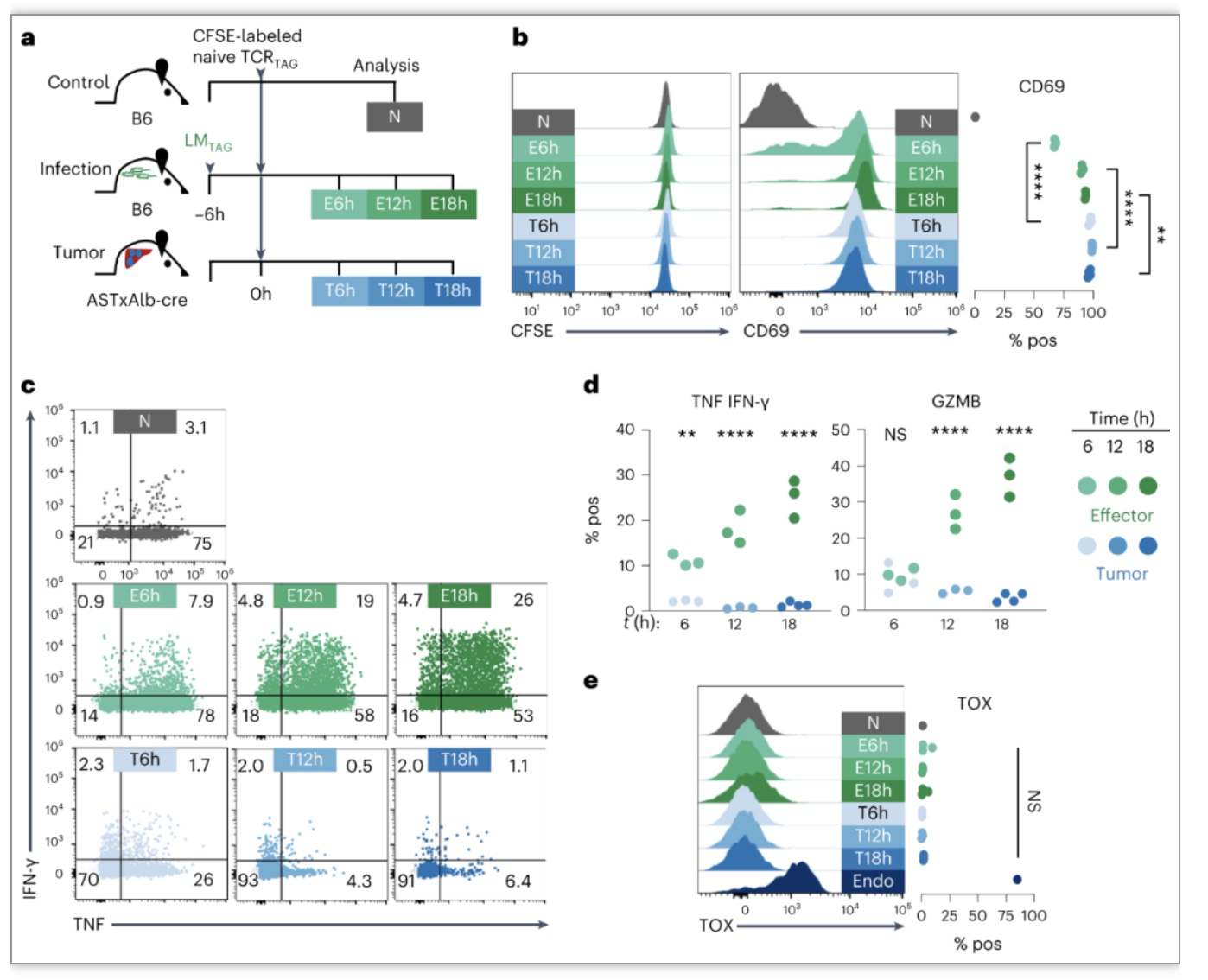
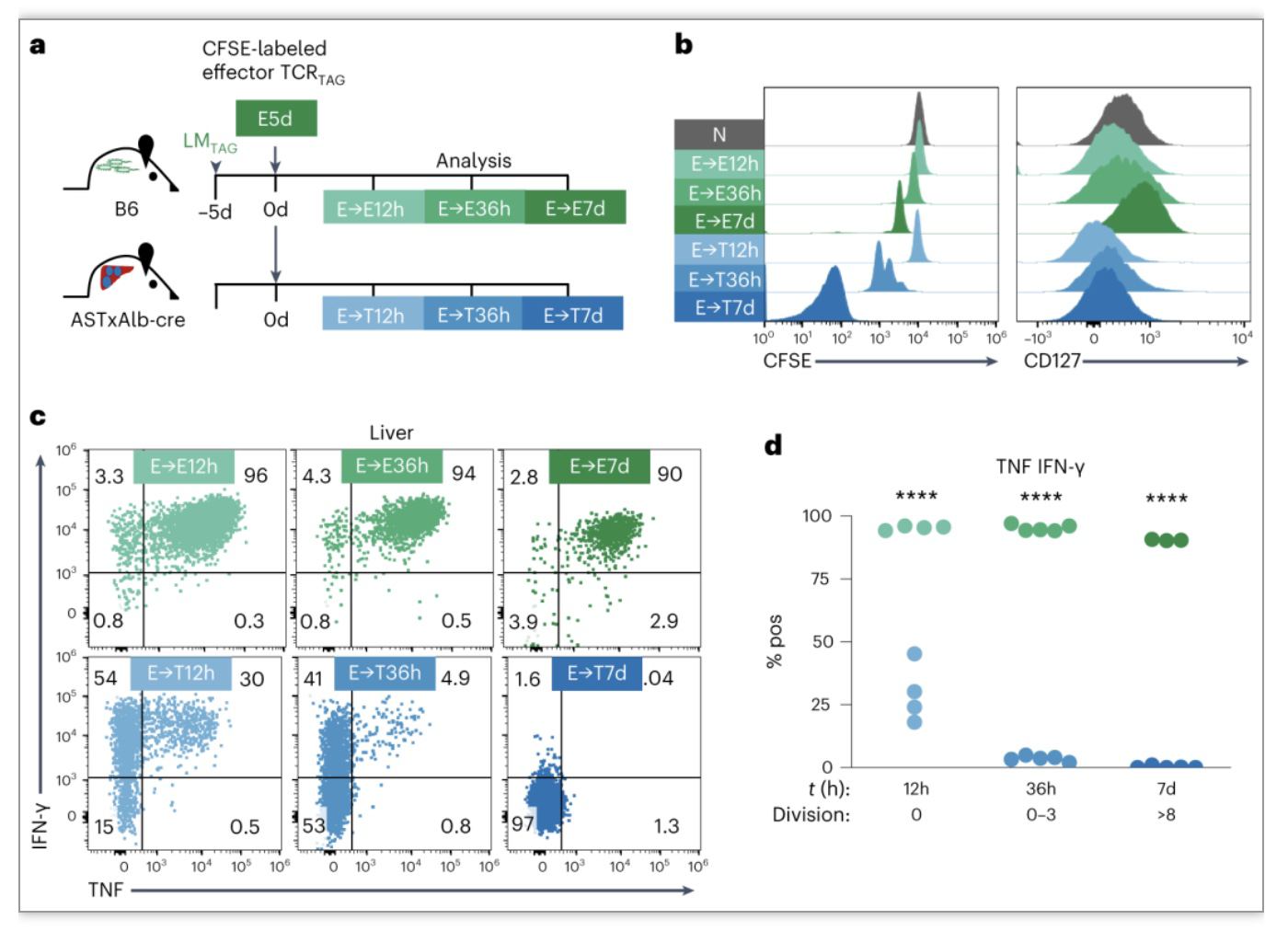
作者通过将表达初始TCR
TAG
的CD8
+
T细胞回输到LM
TAG
感染的小鼠中,并在回输后5天(E5d)收集脾细胞获得TCR
TAG
(E5d)。作者进一步TCR
TAG
(E5d)过继转移到荷瘤 ASTxAlb-Cre (E→T) 小鼠或同时间段的LM
TAG
感染(E → E)的小鼠中,并在回输后12h、36h和7天重新分离TCR
TAG
进行分析。E→E组CD8
+
T细胞具有正常产生效应细胞因子TNF/IFN-γ能力,而
E→T
组C
D8
+
T
细胞转移后具有增殖能力但1
2
h产生细胞因子能力受损,7天完全丧失
。
图3. 效应CD8
+
T细胞在肿瘤中迅速丧失功能
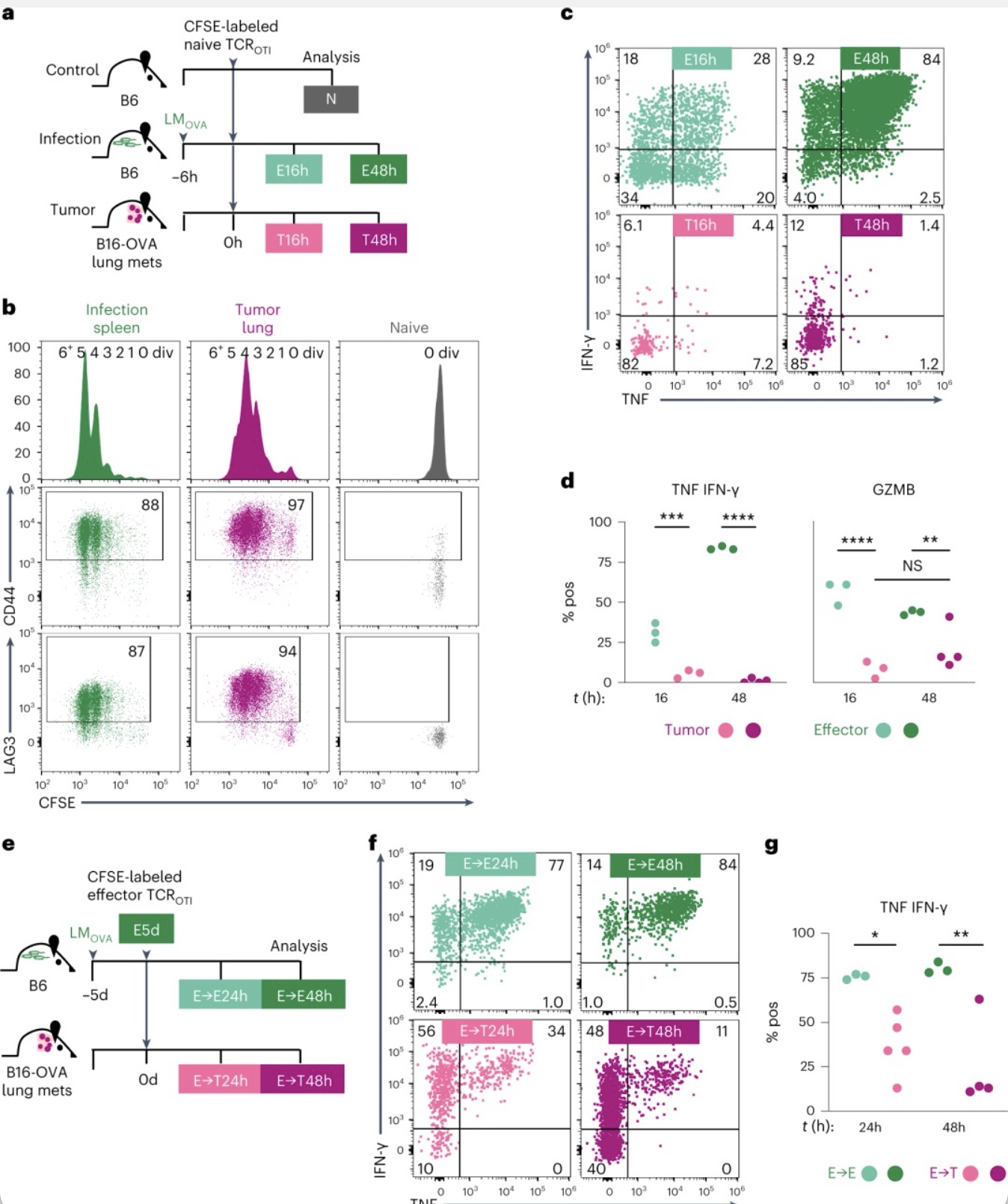
为了判断肿瘤特异性CD8
+
T细胞效应功能的丧失是否发生在具有不同抗原特异性的其它肿瘤中,作者进一步在转移性黑色素瘤小鼠模型中验证了该现象。将特异性识别OVA表位的CD8
+
T细胞(TCR
OTI
)过继转移至携带B16-OVA或LM
O
VA
感染的小鼠中,16h和48h后进行分析。实验发现16h后T-TCR
OTI
和E-TCR
OTI
组CD8
+
T细胞表明 CD69和PD-1表达上调。48h时,T-TCR
OTI
和E-TCR
OTI
组CD8
+
T细胞均显著增殖并上调CD44和LAG3。E-TCR
OTI
组CD8
+
T细胞在激活16h产生TNF、IFN-γ和GZMB,并随着时间的推移变得更加强烈。与之形成明显对比T-TCR
OTI
组CD8
+
T细胞虽然能增殖,但不能产生TNF和IFN-γ以及产生极少量的GZMB。接下来作者测试了OVA特异性效应CD8
+
T细胞是否会在患有黑色素瘤肺转移的宿主中丧失功能。与先前肝癌模型方法一致,作者分离LM
OVA
感染小鼠中CFSE标记的表达TCR
OTI
(E5d)的CD8
+
T细胞 ,并将其过继转移到时间匹配的LM
OVA
感染小鼠 (E→ E) 或已建立B16-OVA肺转移的小鼠中(E→T)。实验发现转24小时内,当大多数群体尚未分裂时,E→E-TCR
OTI
组CD8
+
T细胞可显著TNF 和 IFN-γ,而E→T- TCR
OTI
组开始失去产生TNF和IFN-γ的能力。以上实验表明
肿瘤特异性CD8
+
T细胞效应功能障碍的快速发生并不是肝脏微环境或肝脏肿瘤所特有的,而是可能发生在不同组织不同
晚期
肿瘤中
。
图4. 特异性CD8
+
T细胞在转移性黑色素瘤小鼠中强劲增殖,而效应功能受损。
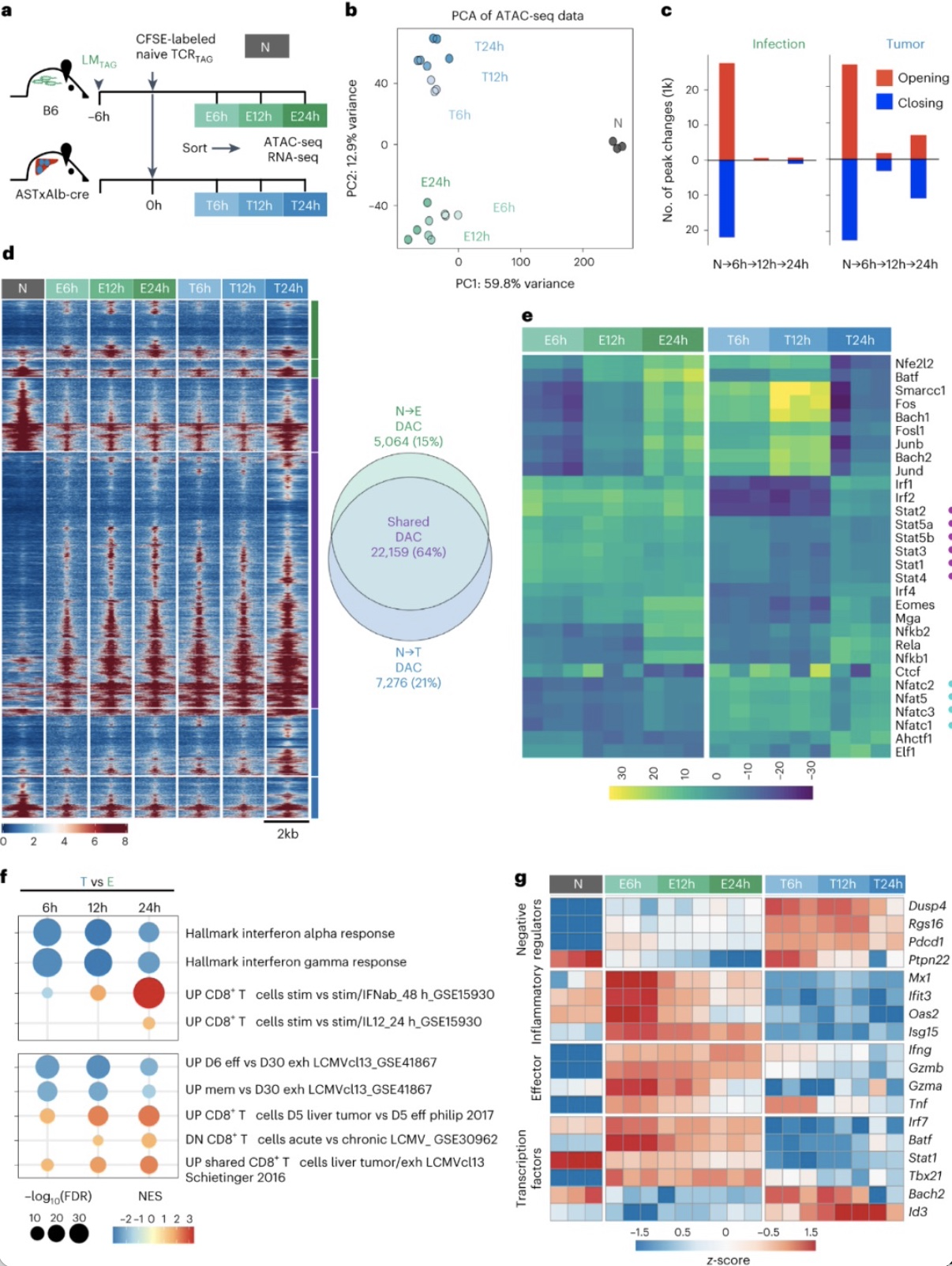
肿瘤特异性CD8
+
T细胞功能障碍发生在分裂前,作者进一步探究了与功能障碍相关的表观遗传重塑是否也发生在细胞分裂之前。作者将6h、12h和24从肝脏肿瘤和脾脏中分选CD8
+
T(TCR
TAG
)细胞进行ATAC-seq和RNA-seq分析。实验发现6h后T-TCR
TAG
和E-TCR
TAG
具有不同的染色质可及性和基因表达谱,并且染色质重塑变化主要发生在激活后的前6h内,而在12h和24h时发生的变化较少。并且与炎症细胞因子诱导的转录因子相关基序(如STAT家族成员)优先富集于早期E-TCR
TAG
组,而在肿瘤特异性CD8
+
T细胞中NFAT(在肿瘤中驱动抑制性受体表达)转录因子家族明显富集。对比T-TCR
TAG
和E-TCR
TAG
组发现早期的E-TCR
TAG
富集了与IFN-α和IFN-γ相关的基因集,与染色质水平STAT基序富集一致,而T-TCR
TAG
组富含没有炎症刺激的T细胞中表达的基因集。以上表明在
感染宿主和荷瘤宿主中激活的
CD8
+
T细胞接受了类似的TCR刺激,然而在荷瘤宿主中激活的CD8
+
T细胞无法接炎症细胞因子信号
。
并且作者发现第6天和第30天时,与病毒慢性感染或较晚耗竭的肿瘤内CD8
+
T细胞相比,早期T-TCR
TAG
组表现出显著的基因表达差异,包括T细胞功能负向调节分子(
Rgs16、Pdcd1、Ptpn22
等)表达增多,炎症相关基因(
Mx1、Isg15
等)、细胞因子和颗粒酶(
Ifng
,
Gzmb
,
Gzma
)及功能分化相关转录因子(
Batf
、
Tbx21
等)表达下降。以上数据表明
早期
与晚期功能失调
的CD8
+
T细胞
相关的表观遗传和转录变化已经在荷瘤宿主中激活后数小时内被诱导
。
图5. 与效应功能障碍相关的表观遗传重塑发生在细胞分裂之前。
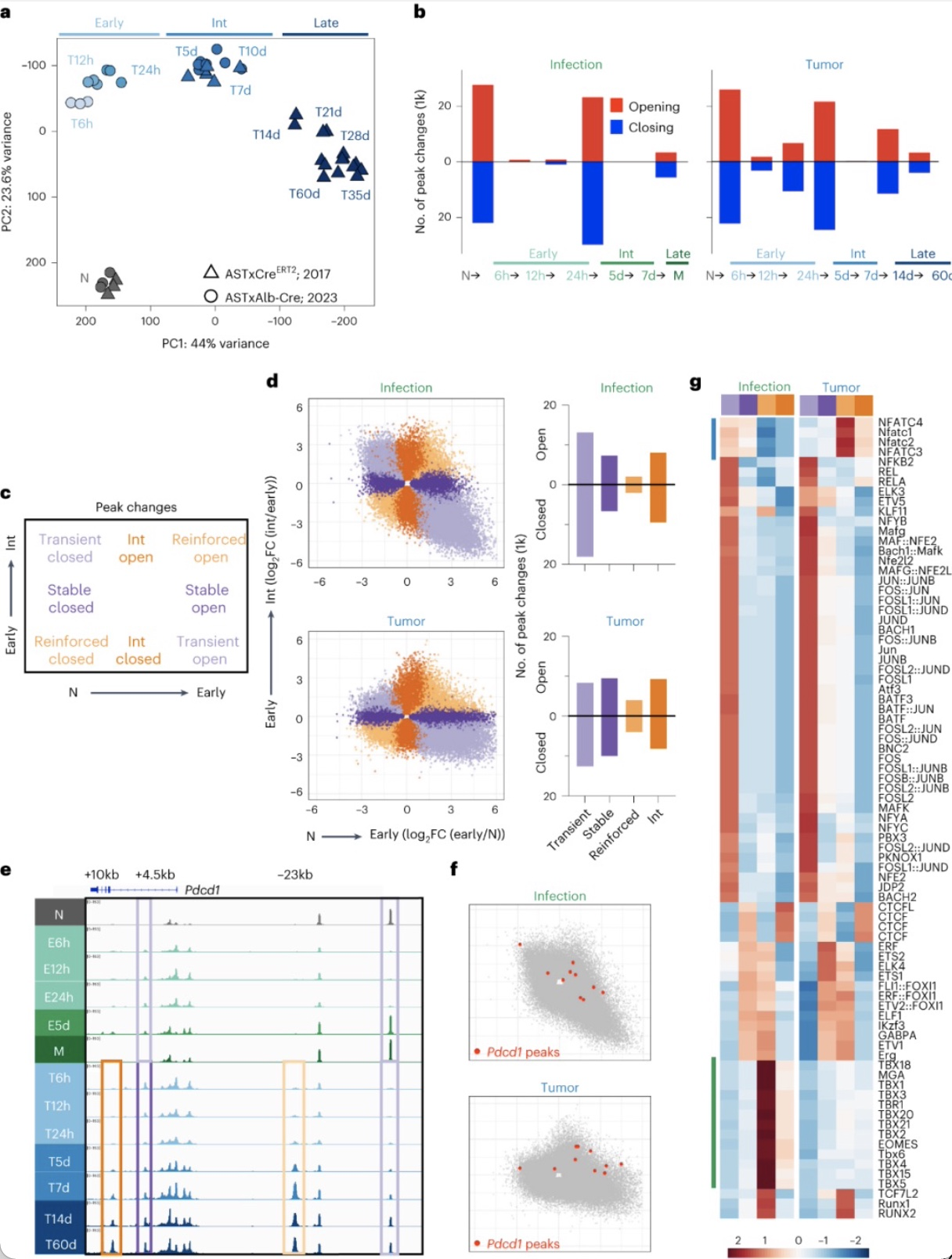
那分裂前功能障碍相关的染色质可及性变化是否会随着时间和持续的抗原暴露而维持或演变?作者将肿瘤特异性CD8
+
T细胞根据肿瘤暴露的持续时间分为三组:早期(6-24小时)、中期(5-10天)和晚期(14-60+天)。作者发现肿瘤特异性CD8
+
T细胞染色质可及性差异最大发生在6小时内,另一轮大的变化发生在24h到5天内,第三轮较小的变化发生在7天到14天之间,此后几乎很少的染色质重塑。作者也研究了早期过渡期间(初始至6h)与中间(int)过渡期间(24h至5天)每个染色质峰可及性的倍数变化如何随时间变化。分析显示随着肿瘤/抗原的持续暴露,肿瘤特异性CD8
+
T细胞中36%的分裂前重塑染色质峰保持稳定,
另外的峰随着时间的推移而增强或在中间过渡时开放。近50%的染色质可及性特征已在肿瘤激活后的前6h内建立,与感染期间的分化形成鲜明对比。
图6. 分裂前肿瘤诱导特异性CD8
+
T细胞染色质重塑随着时间和肿瘤抗原暴露而增强
TCR
TAG
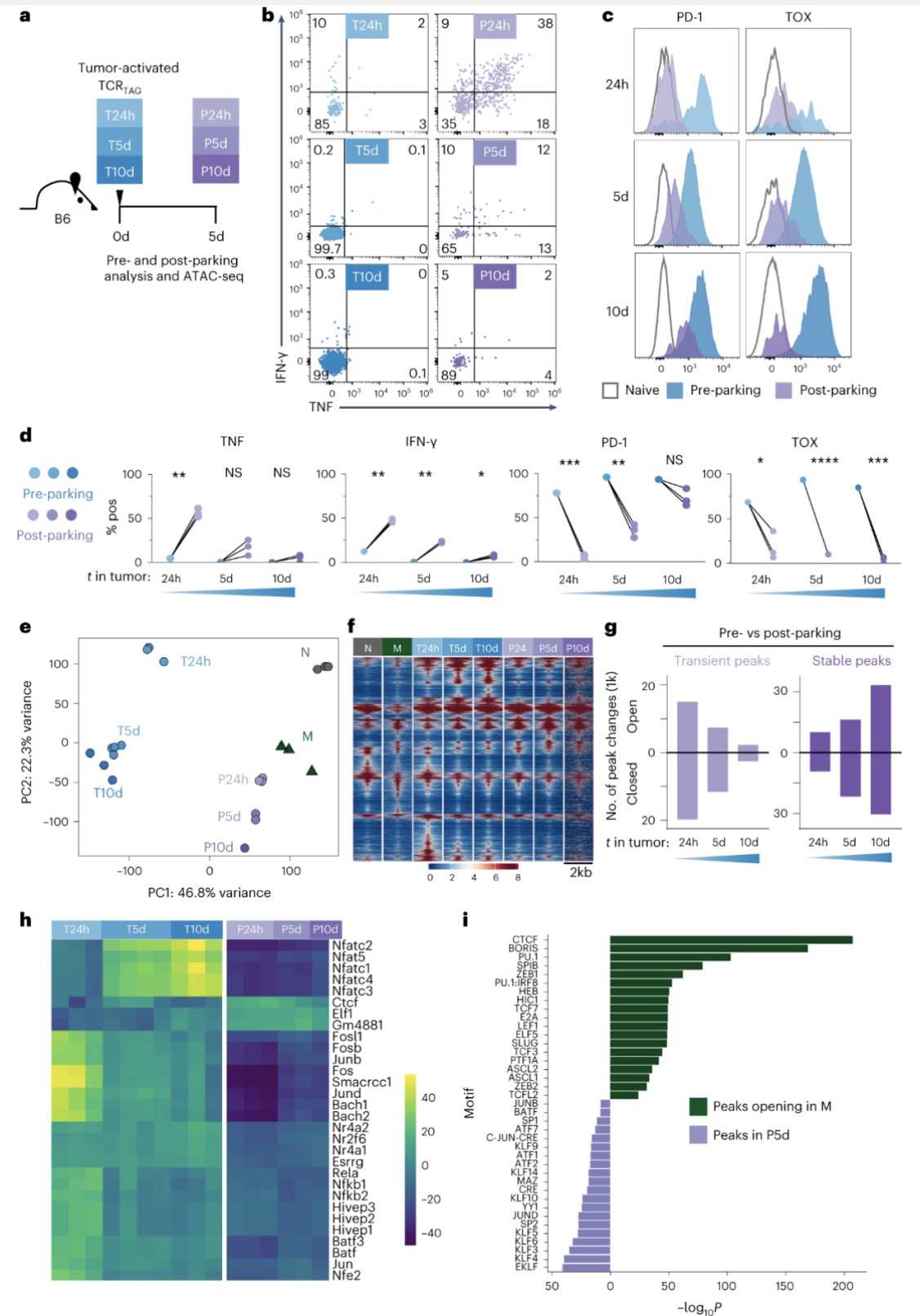
中的许多染色质可及性随着肿瘤抗原暴露的增加而增强,作者猜想如果从肿瘤中去除TCR
TAG
并转移到无肿瘤宿主是否会保留其功能、免疫表型和表观遗传特征。作者在第1、5或10天(T24h、T5d和T10d)从肝肿瘤中重新分离表达TCR
TAG
的CD8
+
T细胞,转移到无肿瘤小鼠(P24h、P5d和P10d)中5天,然后分离进行功能、免疫表型及ATAC-seq分析。转移之前,肿瘤激活的TCR
TAG
不能产生细胞因子。转移到无肿瘤小鼠5天后,P24h下调PD-1表达,但仍有35-40%的CD8
+
T细胞无法产生TNF或IFN-γ。随着肿瘤暴露时间的延长,更多的CD8
+
T细胞效应功能丧失,几乎所有来自P10d的CD8
+
T细胞都无法产生效应细胞因子但保留PD-1表达。染色质可及性分析发现转移到无肿瘤宿主的CD8
+
T细胞与直接分离的或经历过抗原的记忆T细胞的PCA分析明显不同。T24h中的许多CD8
+
T细胞染色质峰值变化是短暂的,转移到无肿瘤宿主中5天后恢复,并且稳定峰值较少,然而随着肿瘤暴露时间的延长,染色质瞬态峰的数量减少,而稳定峰的数量增加。作者T10d将CD8
+
T细胞从肿瘤微环境中分离并转移到非荷瘤小鼠体内,也仍然有相当一部分CD8
+
T细胞无法恢复效应功能,其中与CD8
+
T细胞未能被充分诱导、产生调控分化方向的转录因子密切有关。并且
在肿瘤微环境内停留时间越长,CD8
+
T细胞的表观遗传耗竭“印记”就越明显也越不可逆。
图7. CD8
+
T细胞接触肿瘤抗原的持续时间决定表观遗传功能障碍印记的稳定
文章结论与讨论,启发与展望
综上所述,该研究表明在肿瘤或次级淋巴器官中激活的肿瘤特异性CD8
+
T细胞可能会因肿瘤抗原过多而接收到强烈的TCR信号,从而导致强烈的激活和增殖。然而,肿瘤中缺乏炎症信号导致无法诱导转录因子调节CD8
+
T细胞功能效应分化。荷瘤宿主中持续的肿瘤抗原或TCR刺激或其他负调节信号导致持续的NFAT活性,进一步损害效应功能并诱导负调控T细胞信号和功能的因子表达。
上一篇:
开发低剂量梭曼暴露的磷酸化蛋白质组学灵敏检测技术,表征低剂量化学物质暴露后动物或人体的损伤效应
下一篇:
揭示CLE19-PXL1-SERKs复合物调控花粉发育的分子机制
已经到最底了
技术支持:
库价化学
Copyright © 2024北京螽斯羽生物有限公司 备案号:
京ICP备2023018288号-1