细胞命运可塑性是一个重要的科学问题。利用过表达转录因子或使用化学小分子组合等策略,实现细胞命运转换,即为细胞重编程[1-13]。神奇的是,细胞重编程技术可以将皮肤细胞转变成多能干细胞,将α细胞转变成β细胞,将成纤维细胞转变成神经细胞,近年来受到科学界的广泛关注。由重编程产生的诱导多能干细胞(iPSCs)具备类似于胚胎干细胞的无限自我更新能力和多胚层分化潜能,被广泛应用于疾病建模、药物开发和再生医学。相较于转录因子,小分子具有易于操控、不插入基因组、处理可逆等优点,同时使用纯粹小分子组合的化学重编程也是一种研究细胞命运可塑性的重要体系。然而,目前已报道的化学重编程体系在速率上与传统转录因子重编程存在差距;化学重编程在分子轨迹上与转录因子重编程存在较大差异,说明化学重编程机制上的独特性;目前化学重编程过程的具体分子机制极不清晰,有待深入发掘。
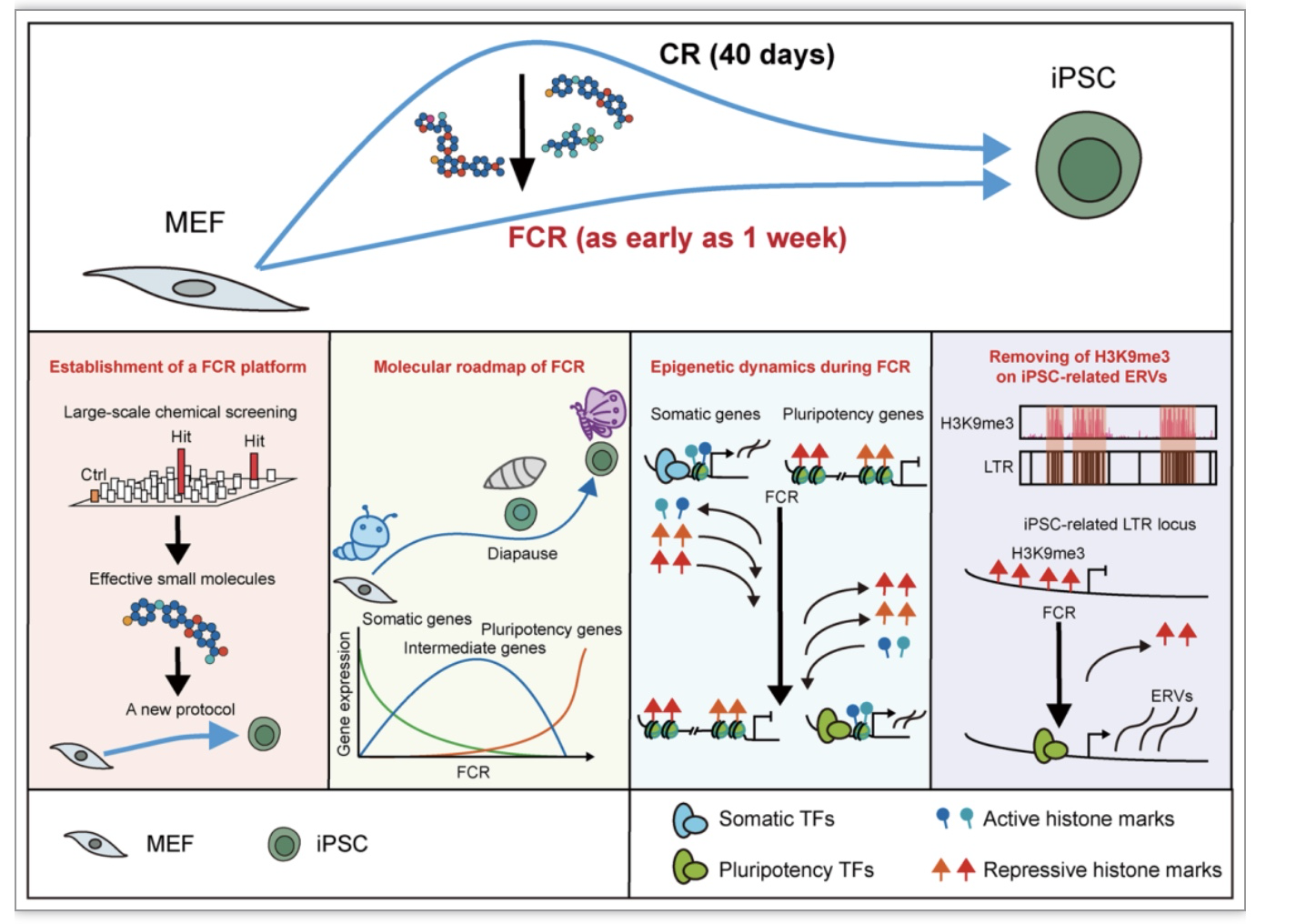
该研究中作者致力于改造小鼠化学重编程体系,通过采取多种策略,筛选了2万多个条件,自主研发创建了FCR,大幅度加快了细胞命运重塑进程,将原先约40天的过程缩短至7-12天。在此基础上,利用多组学测序及整合分析,系统性地展现了FCR过程中的转录与表观遗传动态,并揭示FCR后期经历了一个独特的滞育类似状态。有意思的是,作者还发现了异染色质组蛋白修饰H3K9me3通过调控多能性相关内源性逆转录病毒ERVs从而作为重编程障碍的独特机制。FCR过程中,细胞感受外源信号,克服重重障碍,“破茧成蝶”,华丽转身。
从解决化学重编程周期长、效率低等关键问题出发,作者采取一个与以往研究不同的筛选策略,以最终生成iPSC效率为筛选指标,通过在化学重编程三个阶段分别筛选约7000个小分子,发现了一系列有效小分子可以显著提高重编程效率:第一阶段发现了SR11237、CD3254和CCT129202,第二阶段发现了TSA、TMP269、Citarinostat、 DMH1、AZD9291 mesylate和SGC-CBP30,第三阶段发现了小分子Vc和WS6。进一步测试发现生长因子mLIF、小分子VPA和R406的作用窗口期为2天,提示化学重编程过程可以细分到每2天一个阶段,具备进一步优化空间(图1)。
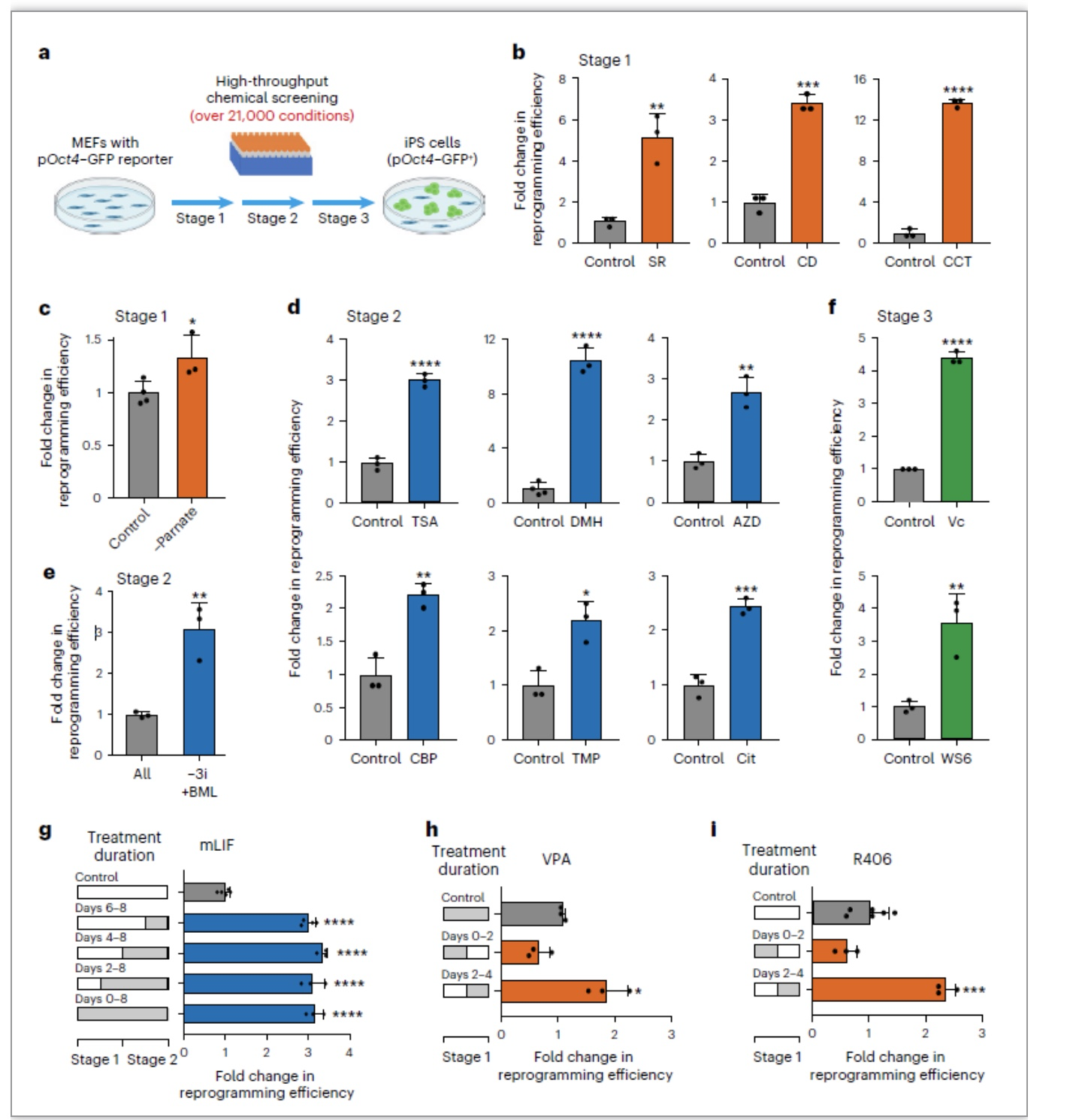
图1. 大规模筛选促进化学重编程的小分子
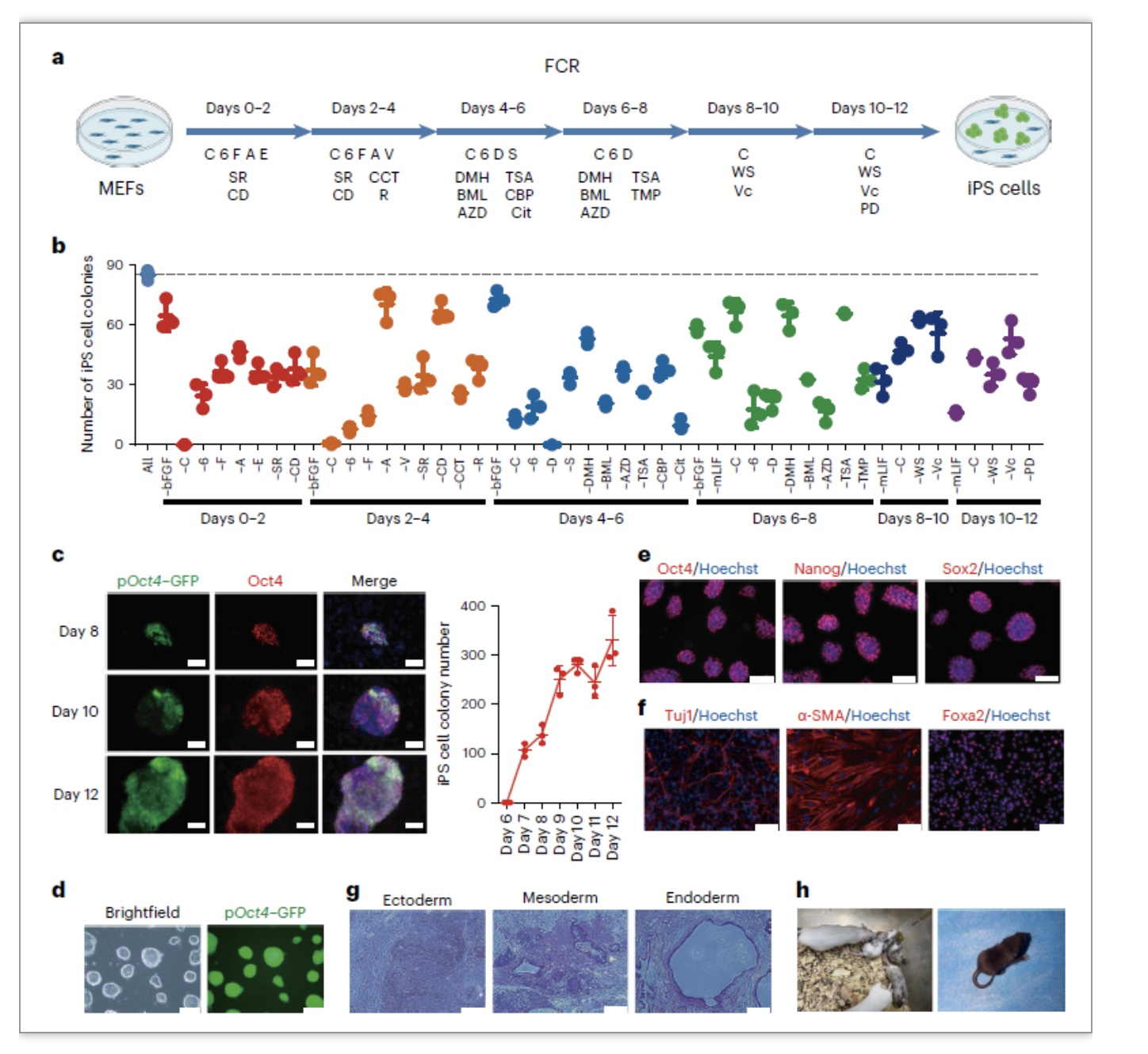
图2. 建立细胞快速化学重编程FCR体系
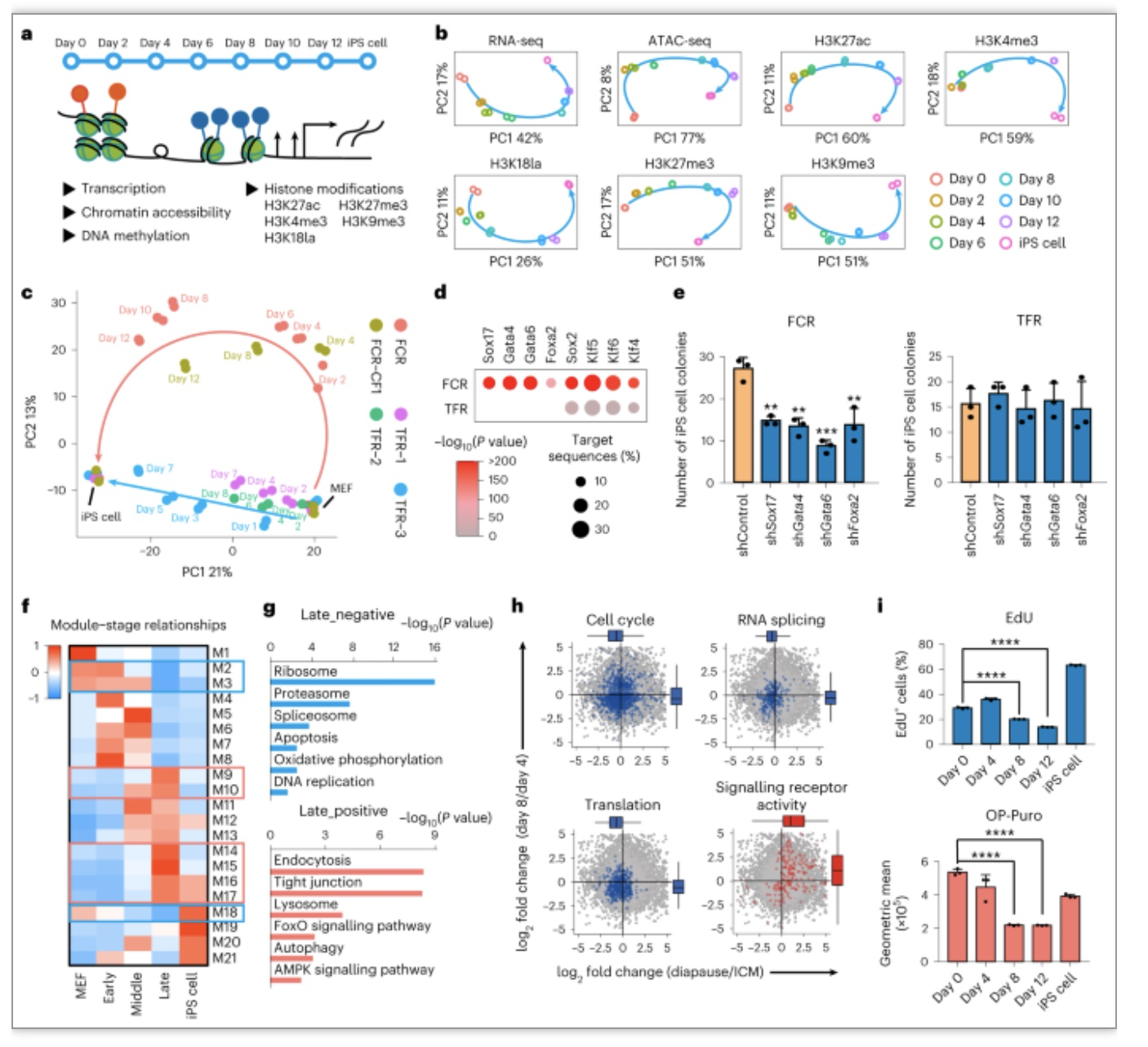
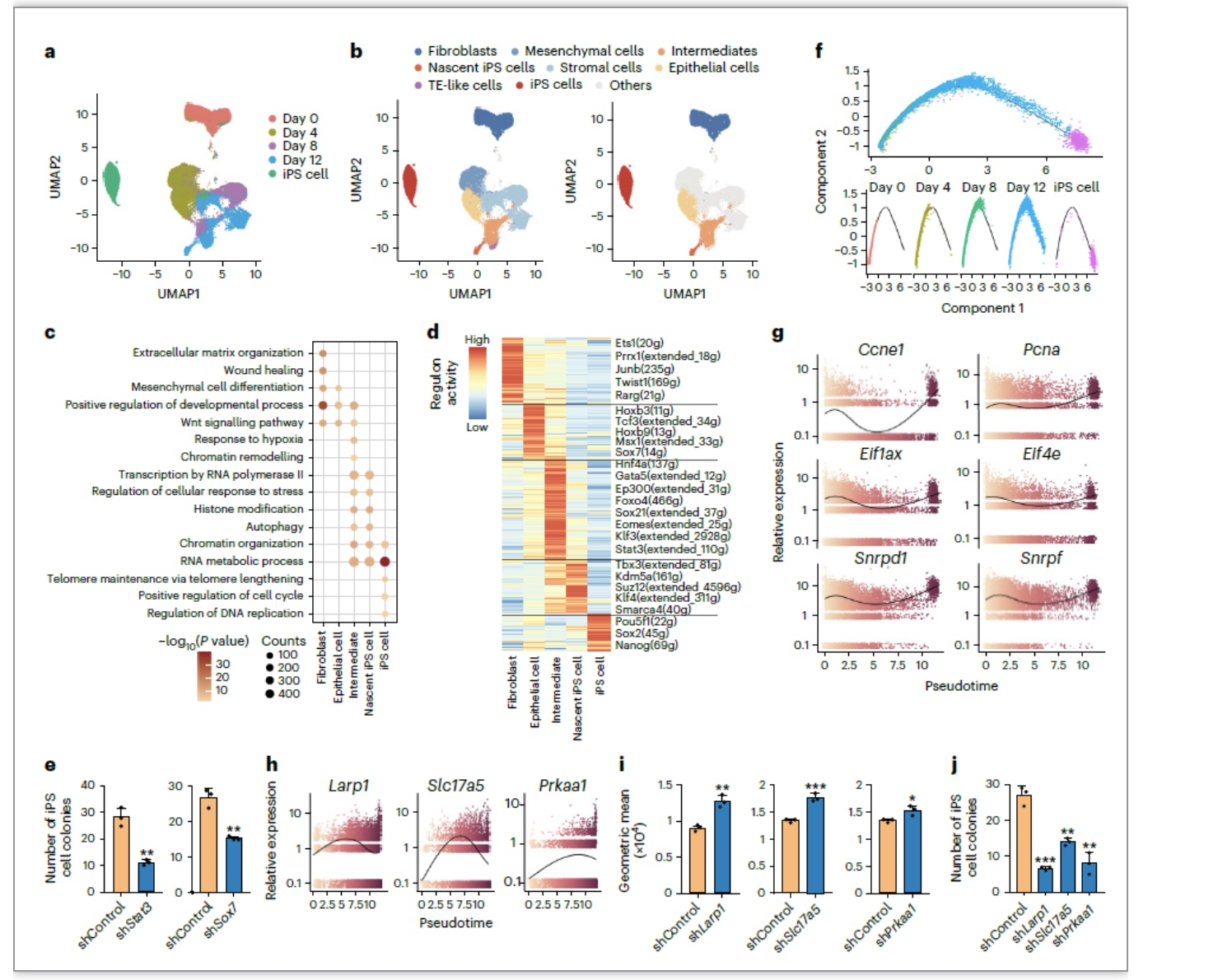
为了排除细胞异质性对RNA-seq结果的干扰,作者在FCR的第0、4、8和12天以及iPSC取样进行了scRNA-seq分析,发现FCR依次经历了成纤维细胞、上皮样细胞、中间态细胞、新生iPSC和iPSC阶段,从而实现细胞命运的逐步重塑。通过SCENIC分析和敲低实验,作者发现并证明了Stat3和Sox7是FCR的重要的中间态转录因子。利用拟时序分析,作者再次证明了滞育相关基因在FCR成功重编程途径后期细胞中的明显下调,并且通过敲低促进细胞进入滞育状态的关键的基因Larp1、Slc17a5、Prkaa1证明抑制滞育类似状态会导致FCR效率降低,说明滞育类似状态是FCR过程中一个重要的中间态,对于细胞内多能性网络的建立起关键作用(图4)。
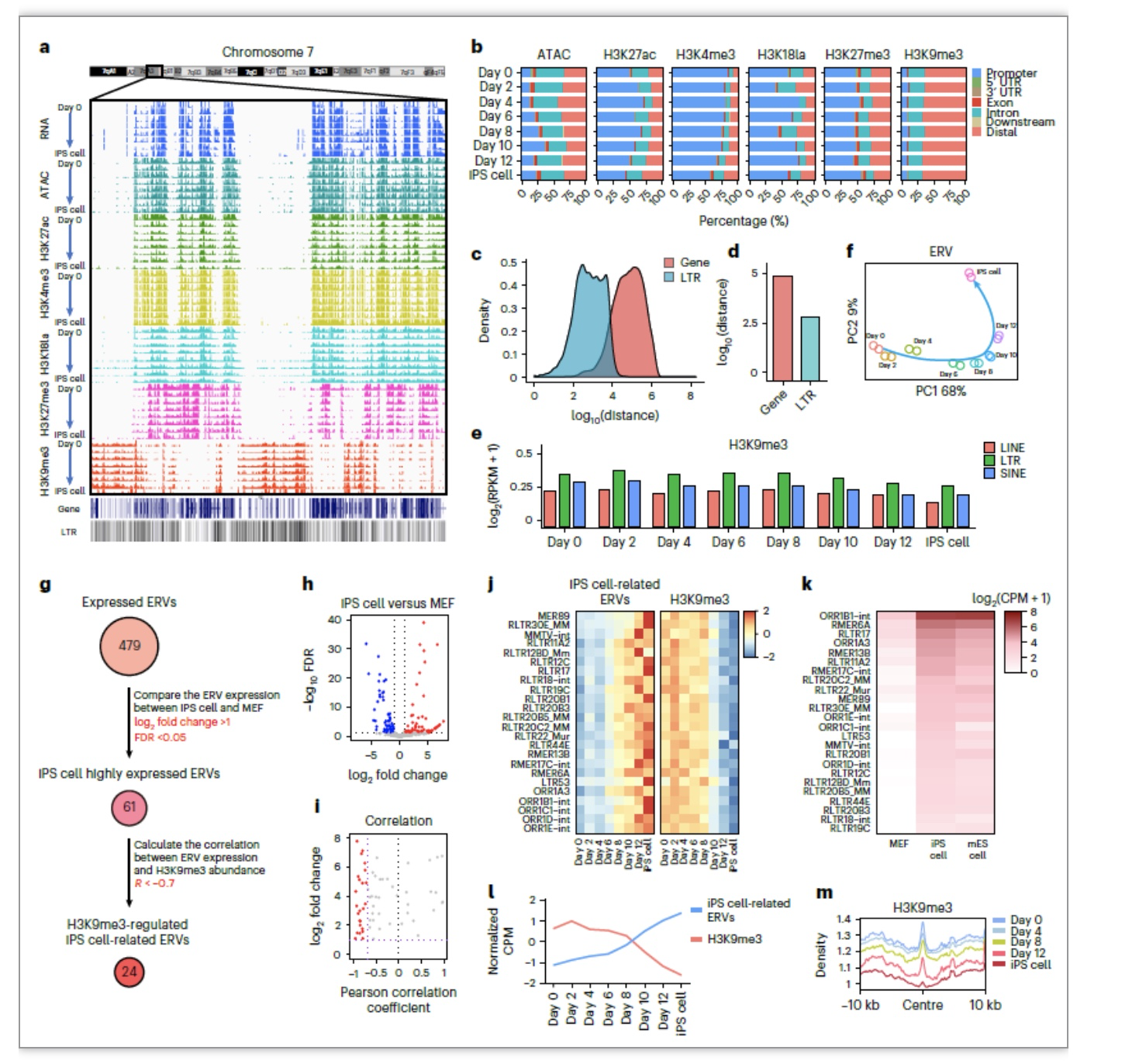
接下来,作者继续抽丝拨茧,整合ATAC-seq和多种组蛋白修饰的CUT&Tag数据进行了联合分析,发现在FCR过程中,动态的染色质可及性位点大部分在FCR中呈现高度动态,伴随着相应组蛋白修饰的动态变化。即使细胞经历滞育类似状态,细胞核内仍发生着剧烈的表观遗传重塑。进一步在全基因组水平上,作者发现异染色质的标志性修饰H3K9me3在FCR过程中都富集于基因组上基因稀疏的区域,这些区域包含大量ERVs。而在FCR过程中,ERVs的表达模式也经历了一个逐步的从MEF向iPSC的动态变化,说明了ERVs可能也会参与并影响重编程过程。H3K9me3已经被证明在多能干细胞中可以调控ERVs,因此,作者筛选找到了24个受H3K9me3调控的iPSC相关的ERVs[18]。iPSC相关的ERVs上有经典Oct4-Sox2-Tcf-Nanog这一多能性转录因子的基序,说明这些ERVs可能参与到多能性网络的构建中。通过敲低和小分子抑制实验,作者证明对H3K9me3甲基转移酶的抑制可以促进FCR,而敲低iPSC相关ERVs导致FCR效率降低。这些结果说明H3K9me3通过抑制iPSC相关的ERVs从而作为FCR的障碍(图5)。