
急性应激通常会导致人类和动物负性情绪的发生,如恐惧、焦虑、抑郁和绝望[1,2]。而这些负面情绪状态的持续存在,可能会引起创伤后应激障碍和重度抑郁症等严重情绪障碍的发生和发展[2,3]。在急性应激、强迫症(obsessive compulsive disorder,OCD)或自闭症(autism spectrum disorder,ASD)等精神疾病模型中,啮齿类动物表现为长时间重复刻板的自我梳理行为(self-grooming),这种行为可以被GABA受体激动剂和5-羟色胺再摄取抑制剂等抗精神病类药物所缓解[4-7]。以往研究显示,丘脑底核(the subthalamic nucleus,STN)的病理生理改变与帕金森病(Parkinson’s disease,PD)的焦虑、抑郁等情绪密切相关[8-10]。在OCD和ASD的治疗中,STN深部脑刺激术或损伤也显示出一定的疗效 [11-13],且光遗传学激活啮齿类动物STN神经元能诱导出反复的自我梳理行为[14]。然而,STN是否参与急性应激相关情绪障碍的调控,以及其相关神经环路机制仍不清楚。

1. STN谷氨酸能神经元参与急性应激诱导的自我梳理行为
作者首先给予小鼠两种急性应激刺激:身体束缚和足部电击,发现小鼠自我梳理行为总时间和次数均明显升高(图1a-n)。且c-Fos免疫荧光染色显示,不论是身体束缚还是足部电击引起的急性应激,均能明显激活STN神经元(图1g-p)。在体光纤成像实验显示,小鼠经历身体束缚后STN谷氨酸能神经元钙信号在每次面部梳理或身体梳理的起始阶段均明显升高,但在随意运动时无明显变化(图2a-g)。以上结果表明STN谷氨酸能神经元参与应激诱导的自我梳理行为。
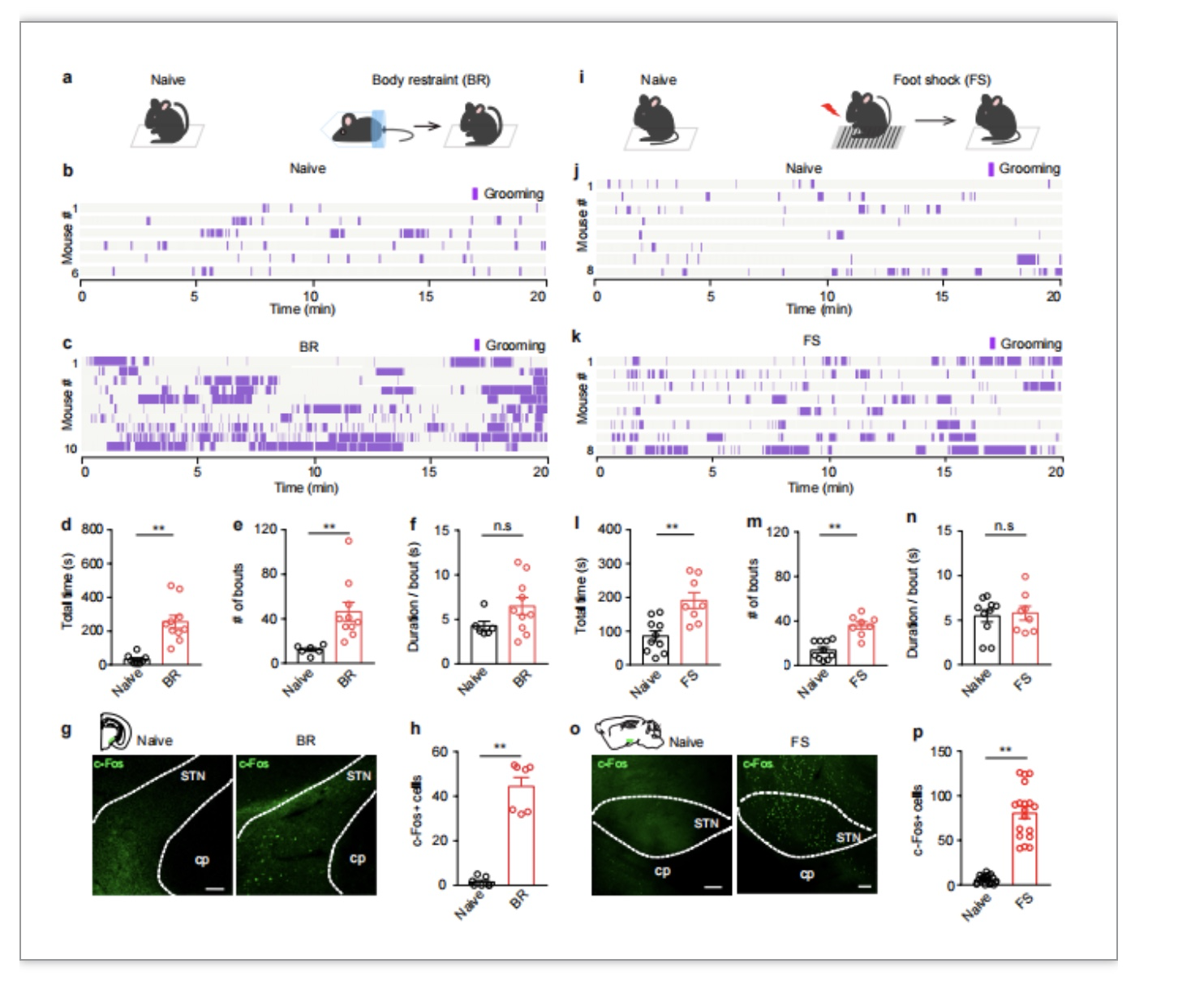
图1. 急性应激导致小鼠自我梳理行为增多,并伴随STN神经元的异常兴奋身体束缚应激。(a)增加自我梳理行为(b, c)的总时长(d)、次数(e),但不影响每次梳理持续的时间(f),同时,会增加STN中c-Fos阳性神经元个数(g,h)。足底电击(i)也能增加自我梳理行为(j, k)的总时长(l)、次数(m),但不影响每次梳理持续的时间(n),也会增加STN中c-Fos阳性神经元个数(o,p)。n.s: not significant. *: P < 0.05. **: P < 0.01。
2. 兴奋STN谷氨酸能神经元可诱导小鼠自我梳理行为
接下来,为了阐明STN谷氨酸能神经元是否调控自我梳理行为,作者采用光遗传学技术激活STN谷氨酸能神经元,发现激活单侧STN谷氨酸能神经元能诱导出重复的自我梳理行为,且与激活频率相关(图2h-p)。与50 Hz相比,10 Hz和20 Hz诱导出重复性自我梳理行为的时间更长,诱导率更高(图2q)。另外,光激活STN谷氨酸能神经元诱导的重复性自我梳理行为以面部梳洗的重复性前肢运动为主,与自发性自我梳理行为比例相似,且激活单侧与双侧STN谷氨酸能神经元诱导的自我梳理行为无明显差异(图2r-t)。结果提示:兴奋STN谷氨酸能神经元可诱导重复性自我梳理行为。
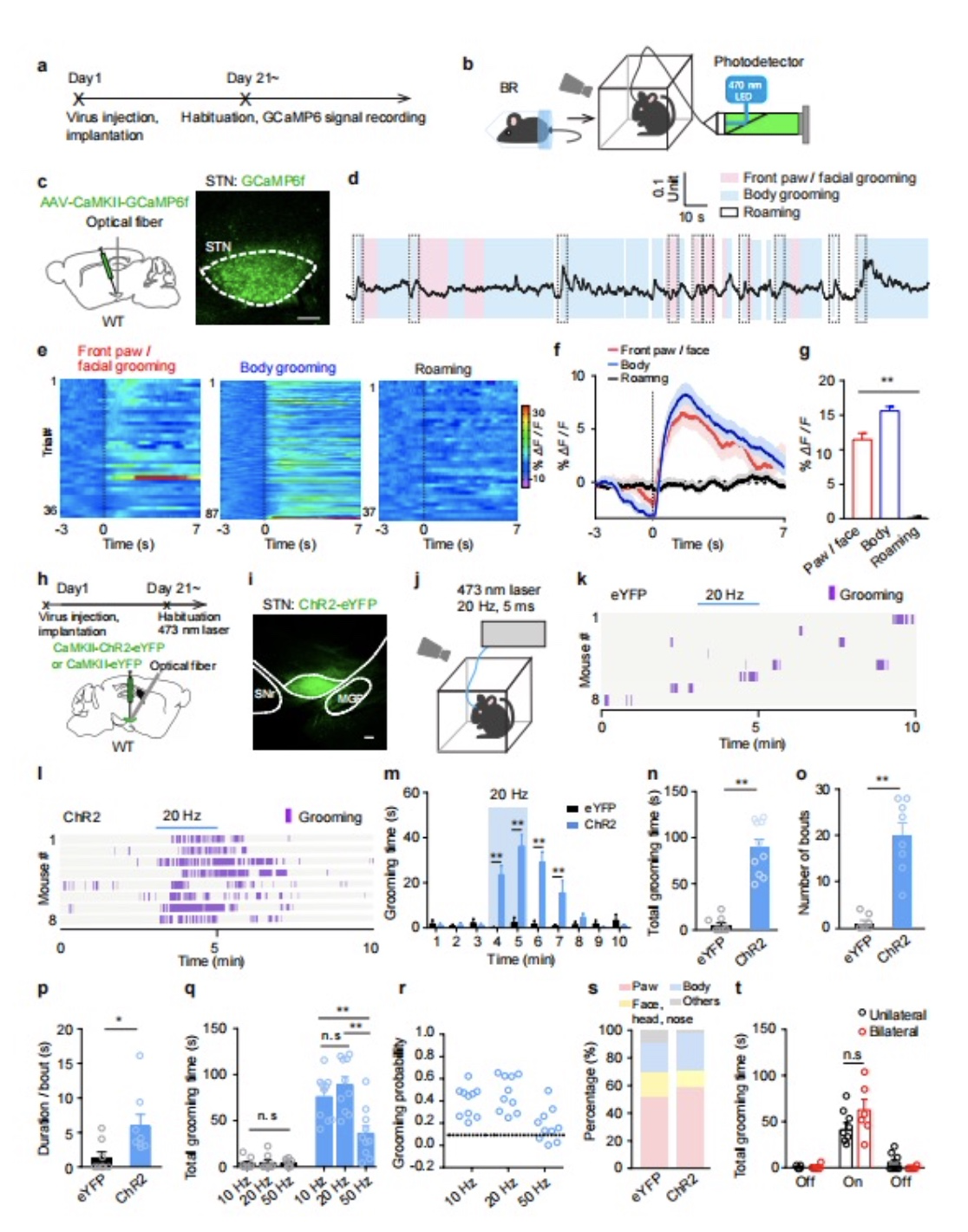
图2. STN神经元参与自我梳理行为的调控。(a-c)用钙离子探针GCaMP6标记STN神经元,在小鼠束缚应激后对STN神经元作光纤成像记录。(e-g)伴随小鼠自我梳理行为,STN神经元GCaMP6荧光增强。用光遗传学技术兴奋STN神经元增强小鼠自我梳理行为(h-p),且有一定的刺激频率依赖性(q-r),但对身体不同部位梳理的行为组成不随刺激频率的变化而变化(s),刺激单侧和双侧STN神经元的作用没有统计学差异(t)。n.s: not significant. *: P < 0.05. **: P < 0.01。
3. STN谷氨酸能神经元通过外侧臂旁核调控自我梳理行为
在明确STN神经元对自我梳理行为的调控作用后,作者进一步探索相关的神经环路机制。以往研究显示,外侧臂旁核(lateral parabrachial nucleus,LPB)接收STN谷氨酸能神经元的调控[15,16],且LPB谷氨酸能神经元可调控负向情绪[17,18],那么STN-LPB谷氨酸能通路是否参与急性应激诱导的自我梳理行为?为回答这个问题,作者首先通过形态学手段解析了STN向LPB的投射。结合Cre依赖的顺向跨突触病毒载体与Ai9报告基因小鼠,发现接受STN谷氨酸能单突触支配的LPB神经元多于内侧臂旁核神经元(图3a-c),结合光遗传学和脑片膜片记录,发现接受STN谷氨酸能单突触支配的LPB神经元绝大多数为谷氨酸能(图3d-h);利用狂犬病病毒揭示支配LPB谷氨酸能神经元的STN神经元多位于STN的内侧(图3i-k)。而无论是光遗传学激活投射至LPB的STN神经元,还是激活STN-LPB通路,均能有效诱导出重复性自我梳理行为(图4a-h,k-r)。且在通过选择性5-HT再摄取抑制剂氟西汀治疗两周后,光激活STN-LPB通路诱导的自我梳理行为明显减少(图4u-x)。除了氟西汀外,作者还发现,同笼小鼠的陪伴也可减少刺激STN-LPB通路诱导的自我梳理行为(见全文图6)。以上结果显示,LPB是STN谷氨酸能神经元调控重复性自我梳理行为的重要下游核团。
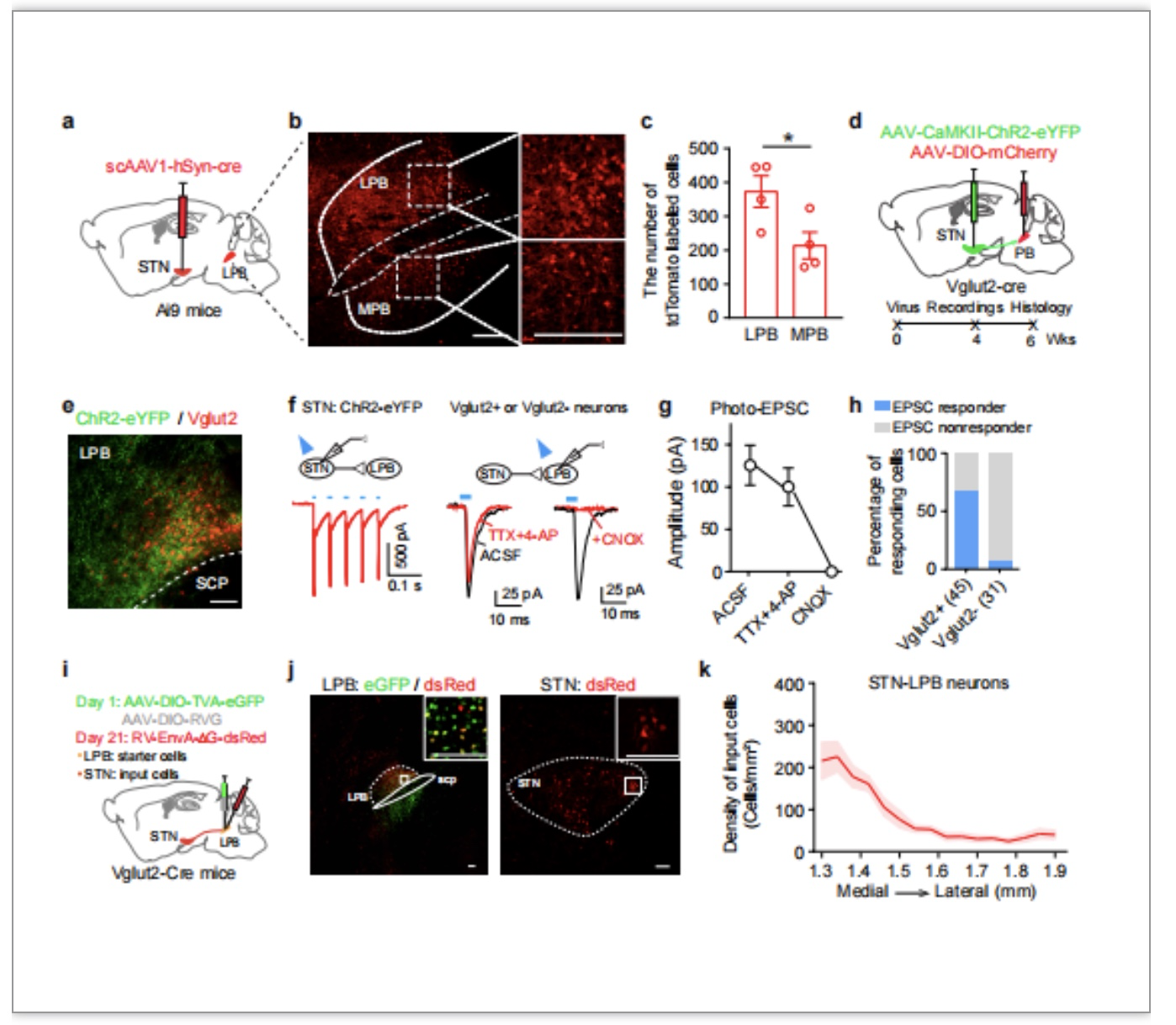
图3. STN通过兴奋性单突触联系支配LPB谷氨酸能神经元。结合顺向跨突触病毒AAV1和Cre报告基因小鼠Ai9(a),证明STN神经元支配臂旁核神经元(b),且被支配的外侧臂旁核(LPB)神经元密度高于内侧臂旁核(MPB)(c)。结合光遗传学技术和脑片膜片钳技术(d-g),发现LPB中接收STN谷氨酸能单突触支配的神经元绝大多数为谷氨酸能(h)。用狂犬病毒做逆向跨突触示踪(i),显示LPB谷氨酸能神经元受内侧STN神经元支配。n.s: not significant. *: P < 0.05. **: P < 0.01。
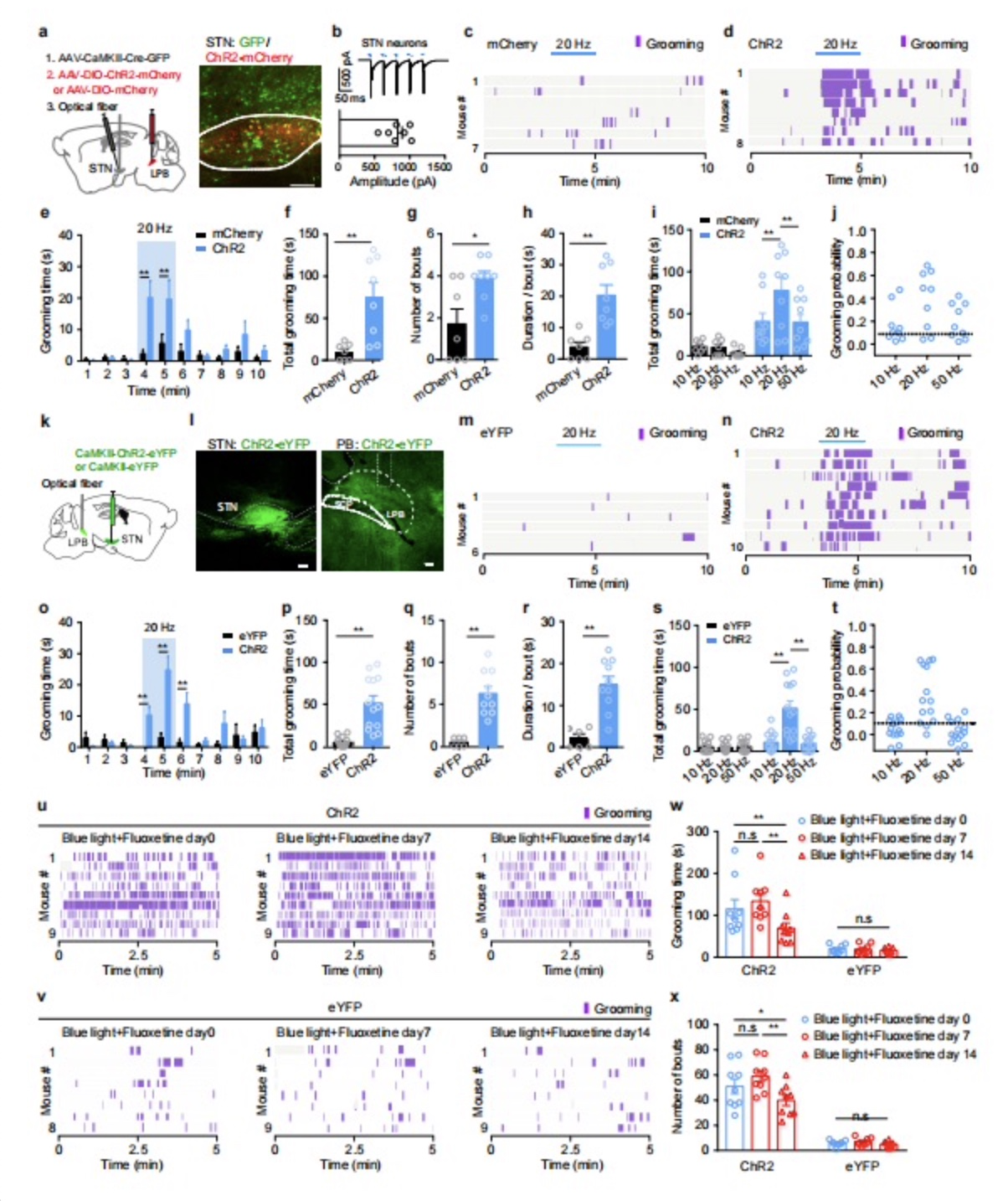
图4. 光遗传学激活STN-LPB通路诱导重复的自我梳理行为。结合逆向示踪和Cre依赖的光遗传学技术标记投射至LPB的STN神经元(a,b)。刺激这些神经元增加了自我梳理行为(c-h),且有时间(e)和频率(i, j)依赖性。用光遗传学技术兴奋STN-LPB投射(k, l)增加了自我梳理行为(m-r),且有且有时间(o)和频率(s, t)依赖性。小鼠饮用氟丁汀(Fluoxetine)14天可缓解兴奋STN-LPB通路诱导的自我梳理行为(u-x)。n.s: not significant. *: P < 0.05. **: P < 0.01。
4. 抑制STN神经元和STN-LPB通路降低急性应激诱导的自我梳理行为
最后,为了明确干预STN神经元和STN-LPB通路是否能缓解急性应激诱导的自我梳理行为,作者利用光遗传学技术抑制STN神经元(图5a-c)和STN-LPB通路(图5 l, m),观察小鼠急性应激的行为学变化(图5d,h,n,r)。结果显示,在接受身体束缚还是足部电击的小鼠中,抑制STN谷氨酸能神经元或STN-LPB谷氨酸能通路,均可使小鼠自我梳理总时间缩短和梳理次数减少(图5e-g,i-k,o-q,s-u)。作者还发现,抑制STN-LPB通路也可缓解D1受体激动剂引起的自我梳理行为(文章补充图)。以上结果显示,STN-LPB通路是急性应激相关自我梳理行为的有效干预靶点。
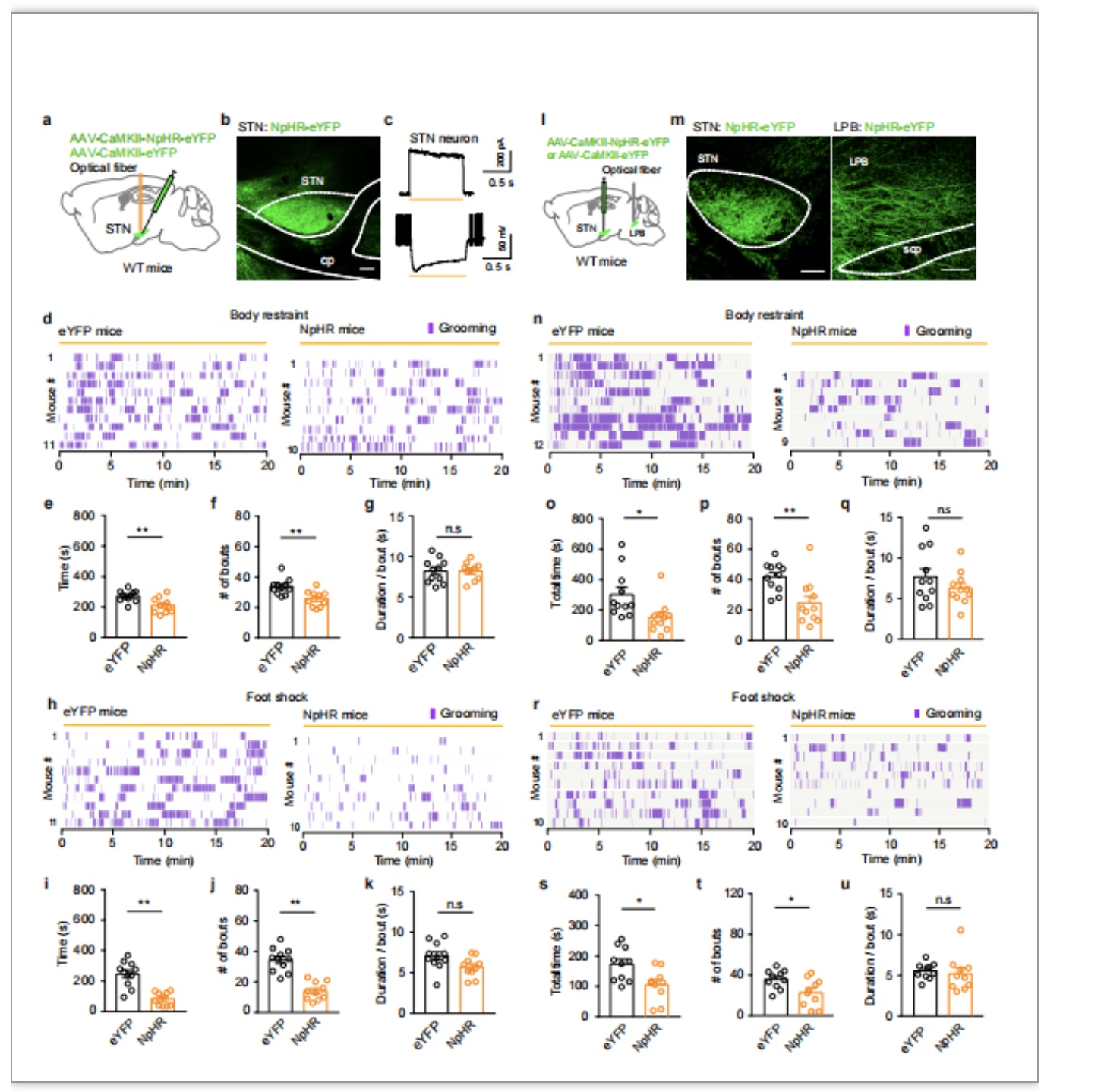
图5. 光遗传学抑制STN-LPB通路降低急性应激诱导的自我梳理行为。用光遗传学技术抑制STN神经元(a-c),可减少身体束缚(d-e)和足底电击(h-k)诱导的自我梳理行为。用光遗传学技术抑制STN-LPB通路(l,m),可减少身体束缚(n-q)和足底电击(r-u)诱导的自我梳理行为。n.s: not significant. *: P < 0.05. **: P < 0.01。
文章结论与讨论,启发与展望
总而言之,本研究采用在体光纤成像、光遗传学、脑片膜片钳方法,结合行为学实验等,阐明了急性应激诱导的自我梳理行为会导致STN和LPB谷氨酸能神经元兴奋性增强;激活STN神经元或STN-LPB谷氨酸能通路均能诱导自我梳理行为;抑制STN神经元或STN-LPB谷氨酸能通路均能抑制急性应激诱导的自我梳理行为。由于STN-LPB通路诱导的自我梳理行为可被氟西汀和同伴缓解,而抑制STN-LPB通路也可缓解D1受体激动剂诱导的自我梳理行为,这些都说明STN-LPB通路调节啮齿类动物情绪障碍相关的自我梳理行为。这些成果提示STN-LPB通路可能是治疗精神疾病相关的重复刻板行为的潜在靶点。
