
卫星RNA由于其缺乏编码复制酶和衣壳蛋白的能力,不能独立复制,因此不能独立存在,需要借助辅助病毒编码的复制酶完成复制,并在寄主蛋白帮助下完成生命循环,属于一类特殊的生物。目前,植物或动物病毒的卫星RNA已有大量研究,显示其一般与辅助病毒没有或仅有很短的同源序列,而且包被在由病毒外壳蛋白构成的病毒粒体里面,然而有关真菌病毒的卫星RNA研究较少。本研究针对一种真菌弱毒病毒卫星RNA开展研究,发现了一类特殊的卫星RNA,分析了其对寄主病毒的影响和生物学特性。

研究人员首先提取了携毒菌株AH1-1中的双链RNA,通过S1核酸酶和DNA酶Ⅰ的消化,共发现三条双链RNA,根据片段长度,自大到小命名为dsRNA1~dsRNA3,无毒对照菌株TP-2-2W进行相同处理,未发现这三条双链RNA(图1B)。进一步的RNA酶A消化显示,在低盐浓度下,这三条双链RNA均被消化,进一步说明了这些片段是双链RNA(图1D)。
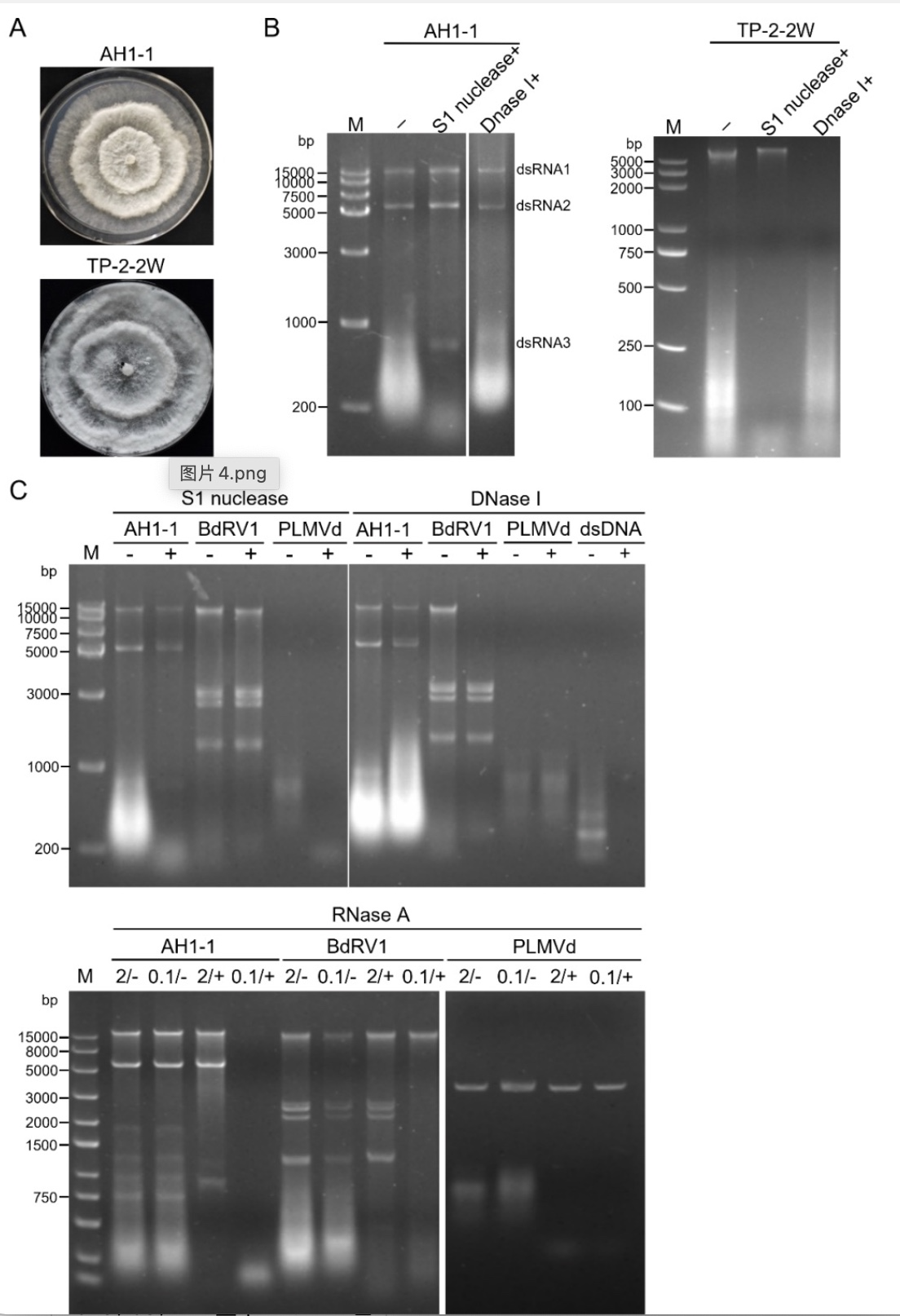
图1 真菌的形态,核酸提取及酶消化
为了进一步研究该病毒的分类地位和基因组结构,研究人员对三条双链RNA进行全长基因组克隆发现,dsRNA1全长10316个碱基对,5′-非编码区有512个碱基对,3′-非编码区有714个碱基对,并在3′-末端有6个腺嘌呤组成的多聚腺苷酸尾巴(poly(A) tail),第513至9602位核苷酸编码一个开放阅读框(ORF),共编码3029个氨基酸(图2B)。根据该ORF构建系统发育树发现该病毒与弱毒病毒科(Hypoviridae)甲型弱毒病毒属(Alphahypovirus)中的多种病毒聚为一支(图2A),故将该病毒命名为无花果拟盘多毛孢弱毒病毒1(Pestalotiopsis fici hypovirus 1,PfHV1)。根据该病毒科共有特性推测,PfHV1基因组由单链RNA组成,双链RNA为其复制中间体。
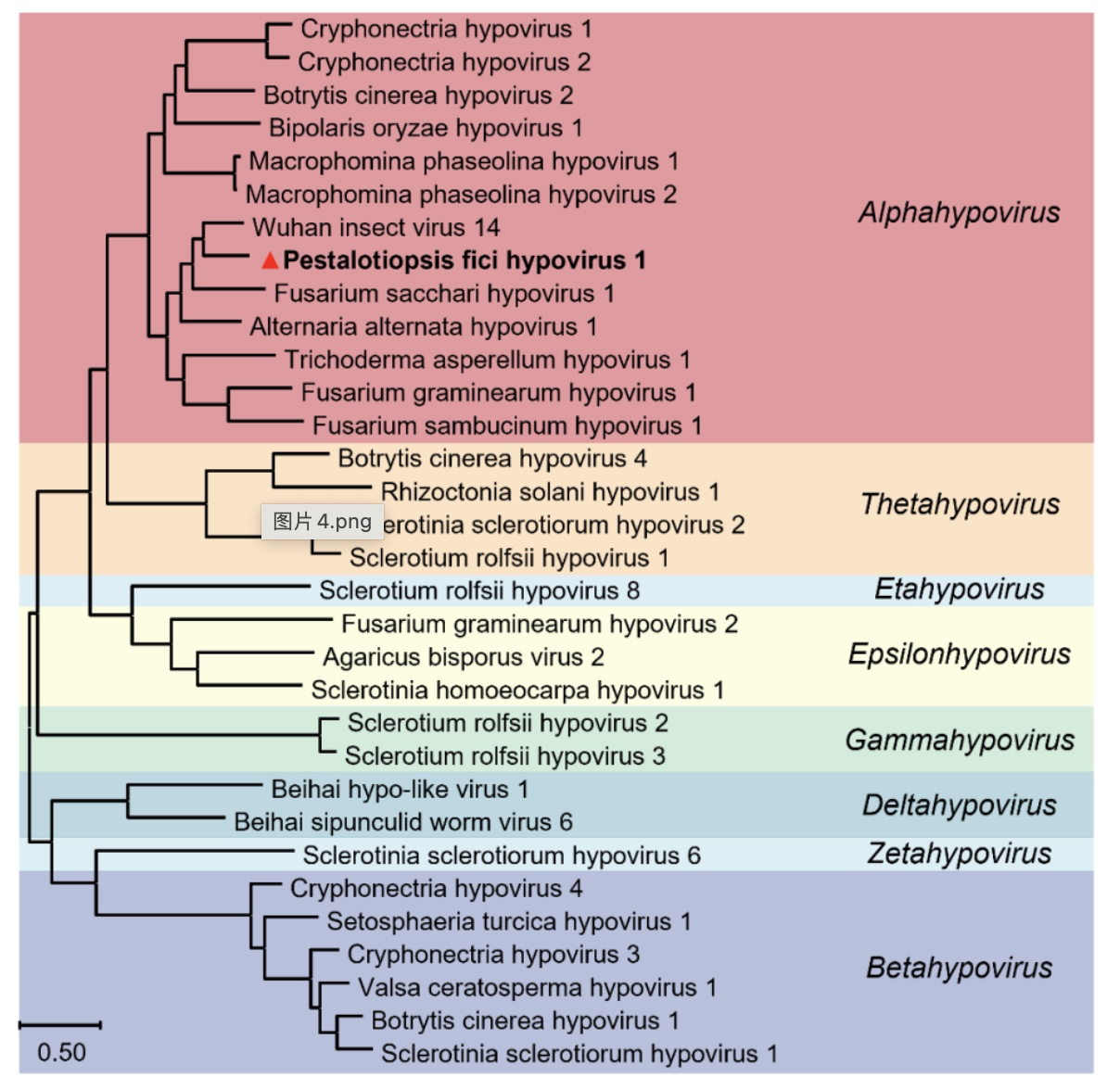
图2 PfHV1的系统发育分析和基因组结构
DsRNA2是dsRNA1的缺陷型RNA,其共有5段序列与dsRNA1完全一致(分别位于第4至3939,5988至6225,6220至6508,6512至6770,6774至7562位碱基),dsRNA3是该病毒的类似卫星RNA(Satellite Like-RNA,SatL-RNA),dsRNA3的第1至170位碱基与dsRNA1完全一致(图3A),且二者能够通过蔗糖梯度密度离心共沉淀,在15%蔗糖层有明显富集(图3D)。这与传统的卫星RNA不具有或仅有少部分序列与辅助病毒同源有很大差异,且与整体病毒科和双生病毒科相比,其不被衣壳蛋白包被。在弱毒病毒科中,已有两例报道的卫星RNA,分别是板栗疫病菌弱毒病毒3(Cryphonectria hypovirus 3,CHV3)的dsRNA3和dsRNA4,核盘菌弱毒病毒1(Sclerotinia sclerotiorum hypovirus 1,SsHV1)的S-dsRNA,其均已被验证能够编码成熟的蛋白质,而根据SatL-RNA的ORF预测结果推测,其不能够编码成熟的蛋白质。
为了探究SatL-RNA对辅助病毒复制的影响,在携毒菌株中超表达SatL-RNA,发现与野生型菌株相比,超表达菌株中辅助病毒的丰度显著降低(图3E),说明SatL-RNA在寄主中的复制干扰辅助病毒的复制,这可能与二者竞争复制酶和寄主复制资源有关。
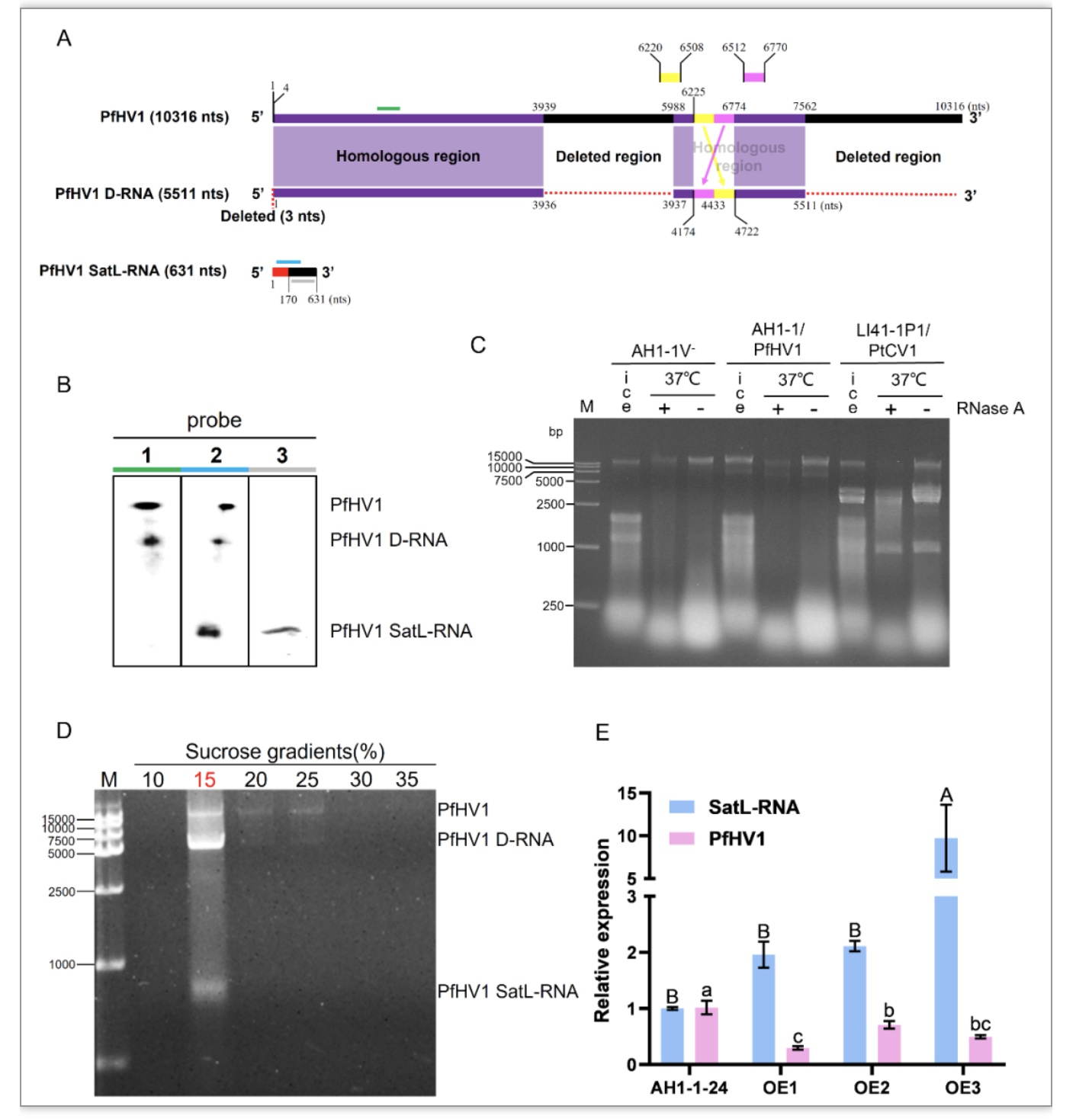
图3 缺陷型RNA和SatL-RNA的鉴定和分析
进一步研究发现,PfHV1对寄主形态、生长速度和致病力无显著影响(图4A~C),与多个无毒菌株相比,携毒菌株AH1-1仍具有较强的致病能力(图4E),说明PfHV1不能发挥弱毒作用。
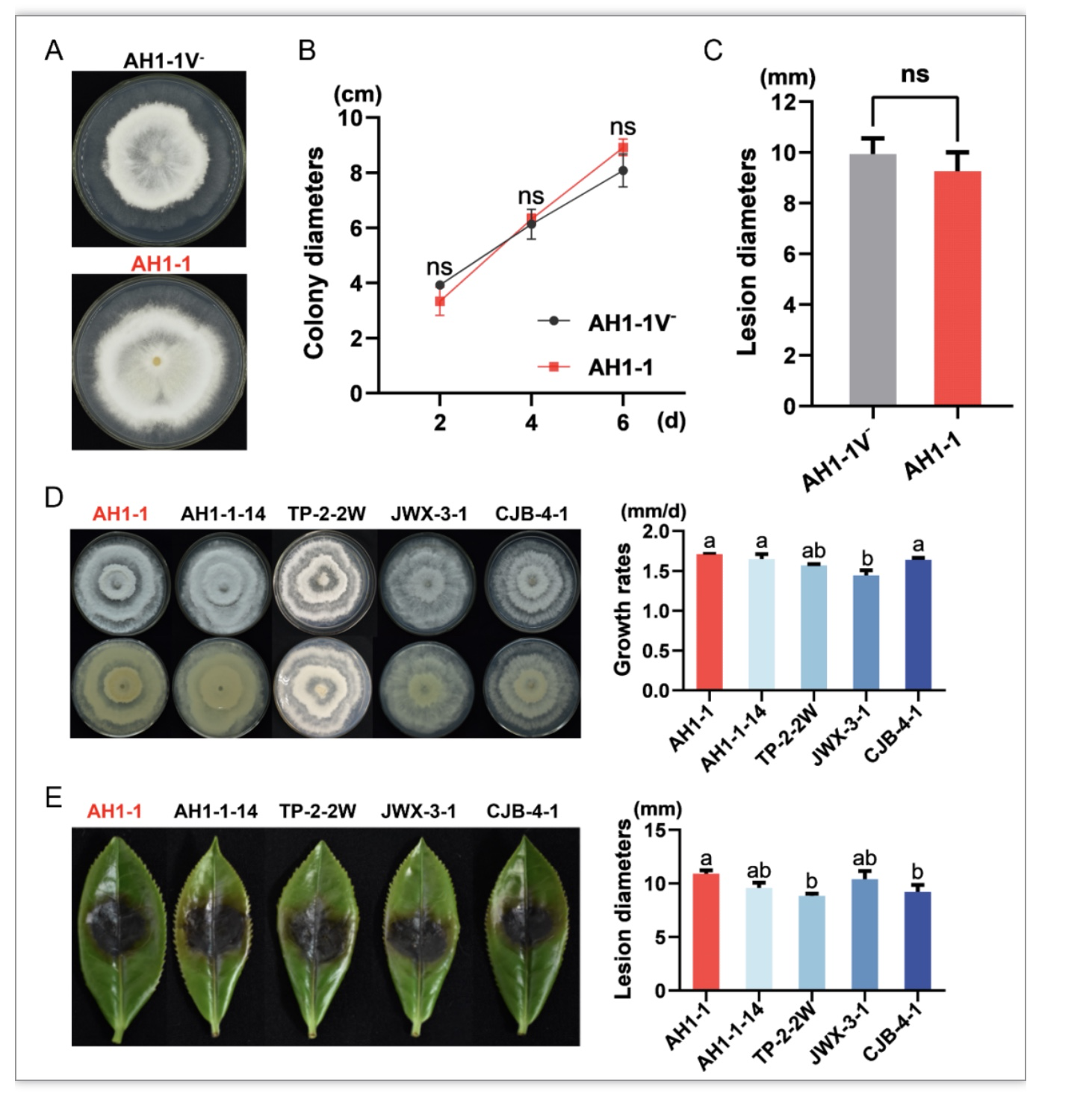
图4 PfHV1侵染对真菌生长速度和致病力的影响
