
豆科植物能够通过与根瘤菌的共生固氮作用获得氮营养,对豆科植物共生固氮机制的研究可以为减少氮肥施用、降低农业温室气体排放提供新的思路。共生固氮发生于豆科植物特有的器官-根瘤。在根瘤组织中,根瘤菌完全进入植物细胞,形成类细胞器结构-共生体(symbiosome)。在一些形成不定型根瘤(有根瘤分生组织存在的根瘤)的豆科植物中,宿主会向共生体传递大量调控信号来促进共生体的终极分化(terminal differentiation)。终极分化状态下,共生体发生内复制,基因组拷贝数显著增多,同时失去繁殖能力。更关键的是,终极分化共生体的基因表达和代谢模式会发生重大变化,固氮还原酶基因被高度激活,从而获得生物固氮能力。是否发生终极分化是自由生长根瘤菌和根瘤细胞共生体的最典型区别,也是共生体能够固氮而自由生长根瘤菌不能固氮的根本原因之一。
终极分化代表着宿主细胞对共生体的精准管理达到了前所未有的新高度。根瘤特异表达的NCR多肽(Nodule Cysteine Rich Peptide)是共生体终极分化的核心调控元件。NCR多肽的序列有着鲜明的特征:初生多肽的N端有着保守的信号肽序列,信号肽被处理后NCR多肽会被运输到共生体中发挥功能;成熟NCR多肽序列中存在4个或6个保守的半胱氨酸位点。豆科植物基因组中NCR多肽的数目往往异常众多,豆科模式植物蒺藜苜蓿基因组一共编码700个左右的NCR多肽。NCR多肽的发现是过去10多年豆科植物共生固氮领域的重大进展之一。但是,学术界对NCR多肽的认识还有很多矛盾之处。例如,在蒺藜苜蓿中,大多数NCR多肽在根瘤固氮区(共生体完成终极分化后进行固氮反应的区域)特异表达,这与NCR多肽调控共生体分化的论点并不一致。固氮区特异表达的NCR多肽是行使何种功能呢?

通过正向遗传学方法,潘怀荣课题组发现了一个蒺藜苜蓿固氮缺失型突变体debino1。debino1突变体只能形成小的白色根瘤(正常根瘤由于大量表达豆血红蛋白呈粉红色),对根瘤细胞进行表型分析发现,debino1突变体发生严重的早衰。通过图位克隆和遗传互补发现,debino1突变体基因组6号染色体上存在着164 Kb的缺失,缺失片段中一个之前没有被报道过的NCR多肽编码基因NCR343是突变体的目的基因。
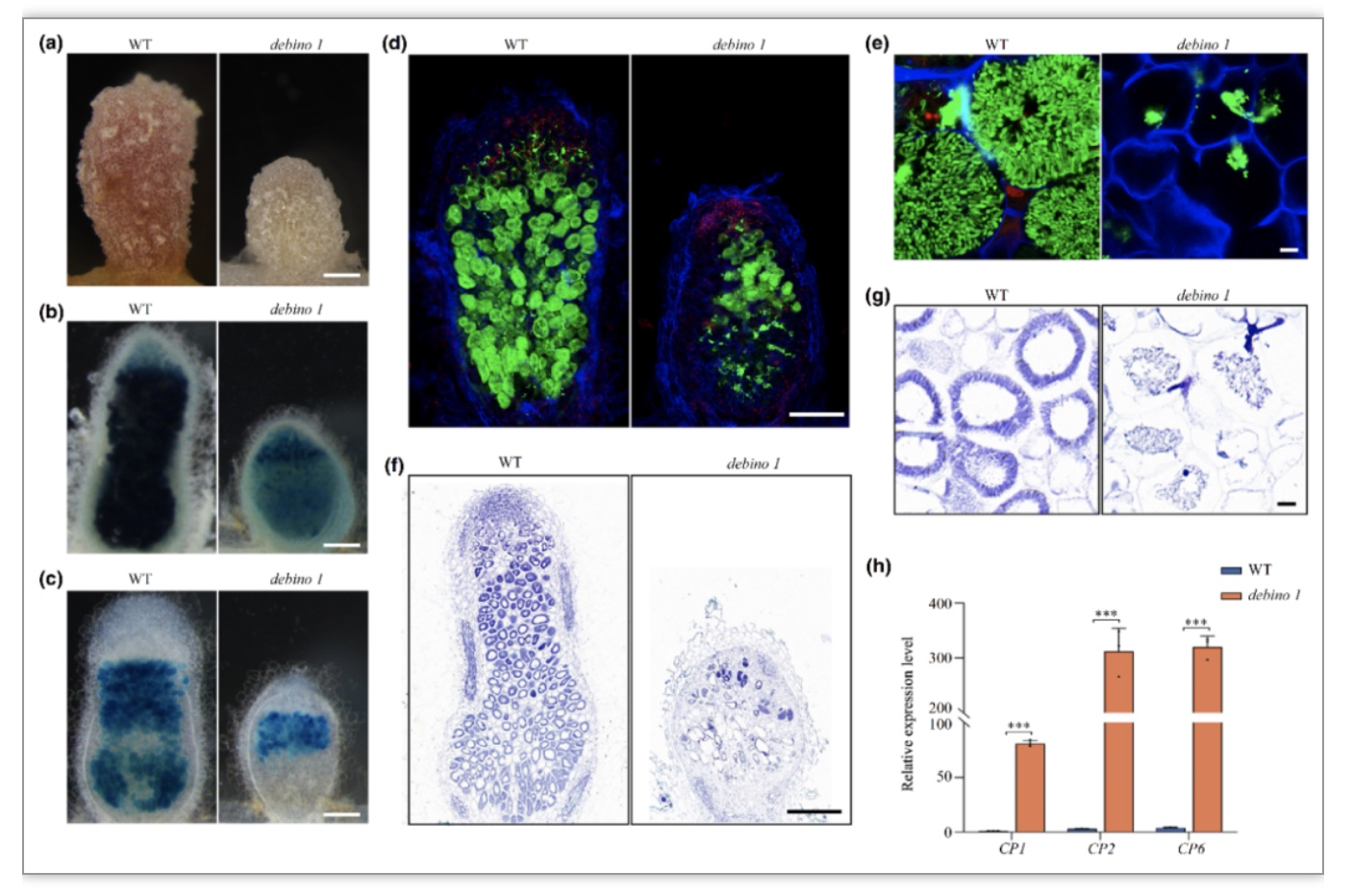
作为一个典型的NCR多肽,NCR343的蛋白序列中含有4个保守的半胱氨酸。表达模式分析发现,NCR343主要在根瘤固氮区表达。通过严格的细胞生物学观察,作者们发现NCR343会被分泌到共生体上,而且可能主要定位在Symbiosome Space(共生体中类菌体与共生体膜之间的间隔区域)中。
为了探究NCR343的功能,作者们对debino1突变体中共生体的分化和存活情况进行了详细的分析。流式细胞仪分析结果表明,在共生体侵染和分化的早期,debino1突变体根瘤细胞中的共生体分化程度和存活状态与野生型非常类似。而在达到终极分化状态后,debino1根瘤细胞中的共生体会迅速发生衰亡。这个结果证明了NCR343是终极分化状态下共生体存活的重要调控蛋白。
为了进一步探究了NCR343调控分化后共生体存活的机制,作者们对根瘤细胞中的共生体进行了分离,并对共生体进行了全基因组RNA-Seq分析,结果表明,debino1共生体中,大量与胁迫应激反应相关的基因被激活表达,甚至还包含一些与氮信号响应相关的基因。同时,借助化学合成的NCR343多肽进行IP-MS实验发现,NCR343主要与一些胁迫应激反应相关的共生体蛋白发生直接互作。这些结果说明,NCR343通过调控共生体的胁迫应激反应促进共生体的存活。
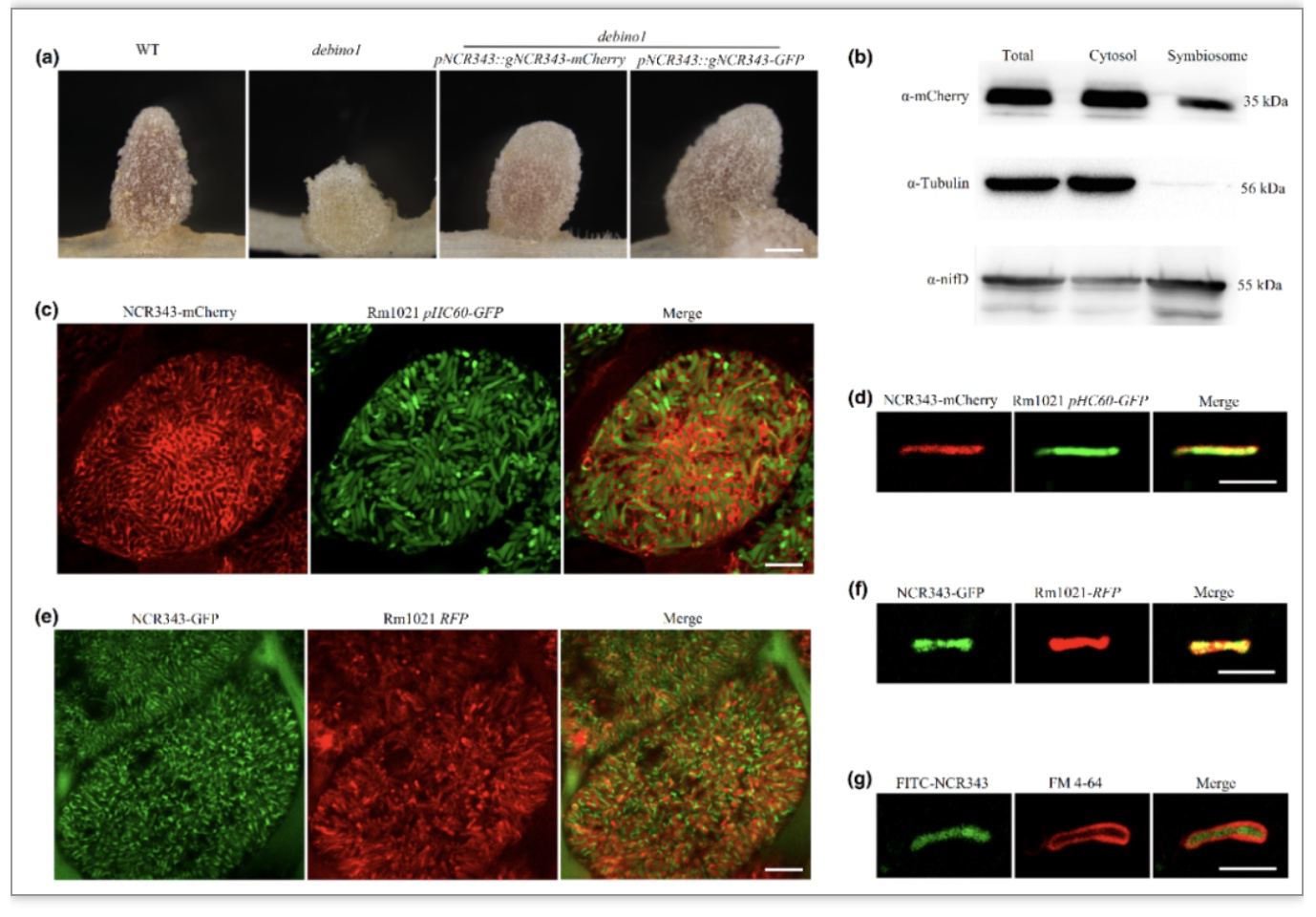
图1:NCR343靶向截茎苜蓿根瘤细胞中的共生体
