
肉毒碱棕榈酰基转移酶1A(CPT1A),催化长链脂肪酸-CoA缀合物的酰基转移到肉碱上,这是线粒体摄取长链脂肪酸类及其随后在线粒体中的β氧化的重要步骤。
Osteopontin
Osteopontin(骨桥蛋白)是一种高度磷酸化的分泌型糖蛋白。是骨组织中发现的第一个细胞外基质蛋白。作为连接最紧密的非胶原蛋白之一,Osteopontin具有调节骨细胞粘附、调节破骨细胞功能和调节基质矿化的功能。
CPT1A实验关键点
CPT1A简介
肉毒碱棕榈酰基转移酶1A(CPT1A),催化长链脂肪酸-CoA缀合物的酰基转移到肉碱上,这是线粒体摄取长链脂肪酸类及其随后在线粒体中的β氧化的重要步骤。
CPT1A的WB检测
CPT1A为多次跨膜蛋白,若煮样后出现条带分子量变大、模糊、拖尾等现象,建议尝试不煮样。
CPT1A定位于线粒体外膜上,若全细胞裂解液检测信号弱,建议尝试提取线粒体组分增强检测信号。
CPT1A存在表达特性,在肝脏组织中表达量较低(如图4-5),若检测靶标蛋白含量较弱的样本(eg:HepG2,肝脏组织),建议增加上样量、降低抗体稀释比例提高检测信号,同时建议增加阳性对照(eg:HAP1、HeLa、SK-OV-3全细胞裂解液,小鼠、大鼠、人肾脏组织裂解液)。
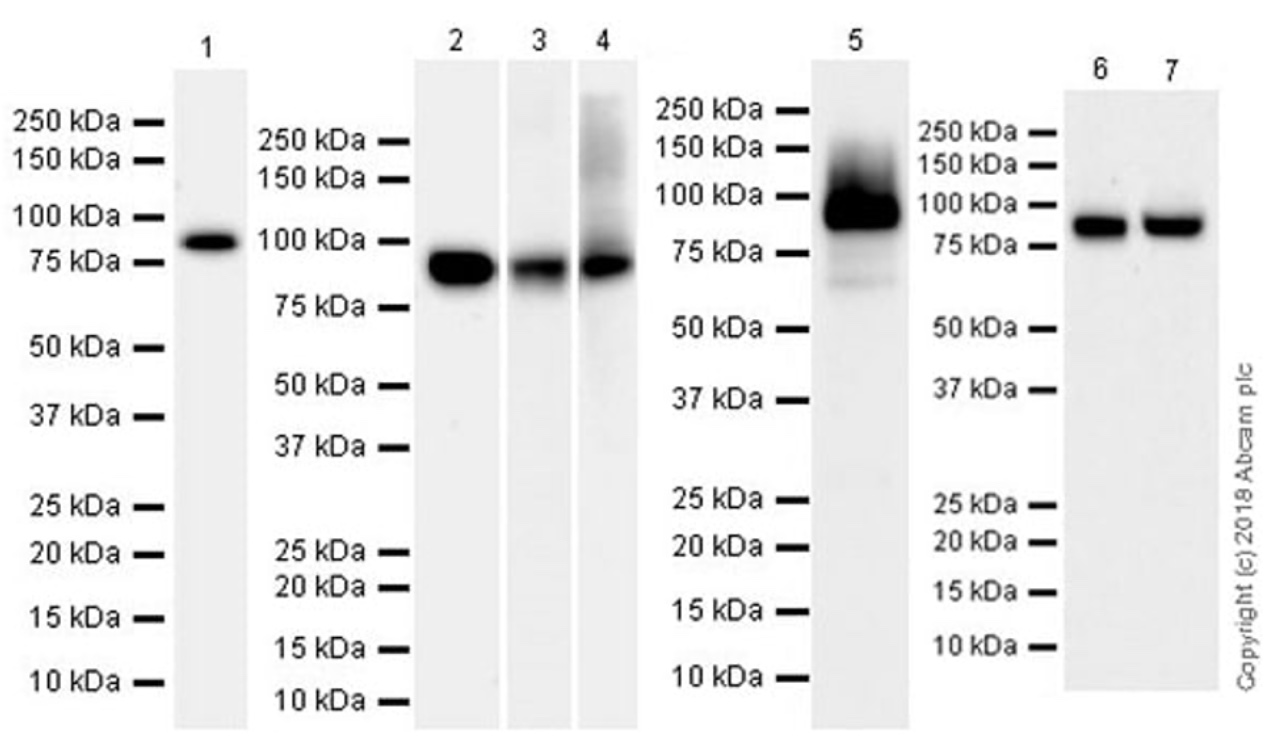
泳道1:20 µg C6全细胞裂解液
泳道2:20 µg MCF7全细胞裂解液
泳道3:20 µg 小鼠肾脏组织裂解液
泳道4:20 µg 大鼠肾脏组织裂解液
泳道5:20 µg 人肾脏组织裂解液
泳道6:20 µg HeLa全细胞裂解液
泳道7:20 µg SK-OV-3全细胞裂解液
预测条带大小:88 kDa
Osteopontin简介
Osteopontin(骨桥蛋白)是一种高度磷酸化的分泌型糖蛋白。是骨组织中发现的第一个细胞外基质蛋白。作为连接最紧密的非胶原蛋白之一,Osteopontin具有调节骨细胞粘附、调节破骨细胞功能和调节基质矿化的功能。
Osteopontin在发生炎症时,可以增强巨噬细胞和T细胞的作用,并可以抑制异位钙化和过度矿化。Osteopontin作为一种多功能的蛋白,在心血管疾病、癌症、糖尿病和肾结石疾病以及炎症、生物矿化、细胞活力和伤口愈合过程中具有重要作用。
实验结果:共聚焦图像显示诱导刺激处理组,LPS (100 ng/ml)处理6小时后,加入BFA (300 ng/ml)处理3小时后,RAW 264.7细胞的细胞质染色增加。
Osteopontin的WB检测
由于Osteopontin存在糖基化修饰和磷酸化等翻译后修饰,可能会在实验中检测到多条蛋白条带。导致实际检测条带大小可能到位于32、45、60、70、90 kDa,与预测值32 kDa可能不符(如图8所示)。
Osteopontin可能会由于交联形成聚集体,可能会在检测到多条条带。建议保留全膜来检测Osteopontin,防止丢失目的条带。
Osteopontin作为一种分泌型蛋白,我们推荐加入BFA处理细胞,防止蛋白分泌指细胞外导致无信号现象。
Osteopontin在不同样本中表达丰度不同,有些样本需要诱导刺激才能够检测到目的蛋白条带(例如加入LPS和BFA处理RAW 264.7细胞才能检测到目的蛋白条带)。推荐使用经过文献验证的实验样本。强烈推荐添加阳性对照(人类肾脏组织、人类尿液等),以确认实验体系没有问题。
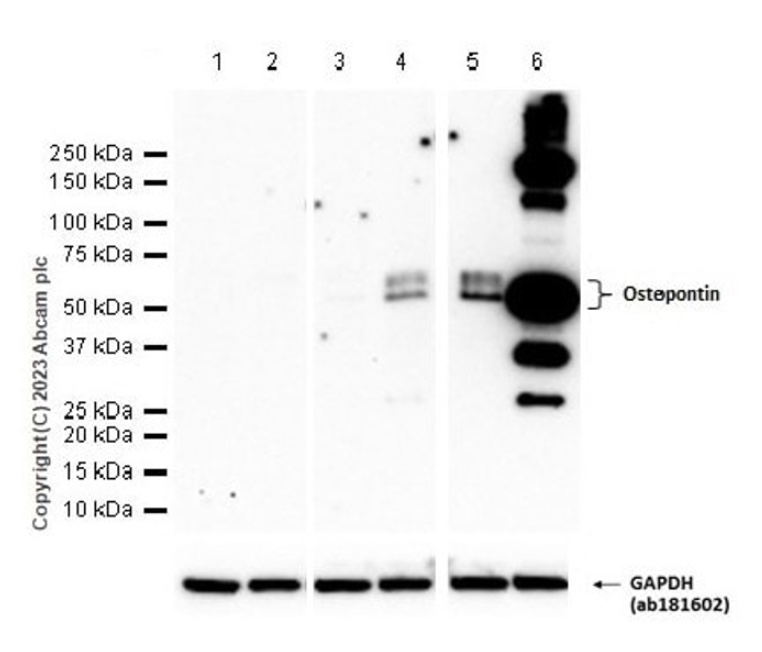
泳道1:小鼠肾组织裂解液
泳道2:小鼠胎盘组织裂解液
泳道3:RAW 264.7全细胞裂解液
泳道4:用100 ng/ml脂多糖(LPS)处理4h的RAW 264.7全细胞裂解液
泳道5:用1000 ng/ml BFA处理3h的RAW 264.7全细胞裂解液
泳道6:用100 ng/ml脂多糖(LPS)处理4h后,再加入1000 ng/ml BFA处理3h的RAW 264.7全细胞裂解液
预测条带大小:35 kDa
实际检测条带大小:60 kDa
样本制备
添加复合蛋白酶抑制剂以避免靶标蛋白降解。
整个样本制备过程中,保持样本置于冰上。
通过Bradford分析、Lowry分析或BCA分析测定样本总蛋白浓度。
至少上样20 μg总蛋白进行电泳。
SREBP1简介
SREBP1(sterol regulatory element-binding protein)是一类转录因子,属于bHLH-zip(basic helix-loop-helix-leucine zipper)家族,能够与甾醇调节元件-1(SRE1)结合。SREBP1信号通路通过一系列负反馈机制调控着细胞内固醇类物质的稳态。
SREBP1合成后以非活性前体的形式存在于内质网。当固醇浓度低时,SREBP1以依赖于COPII的方式从内质网转运到高尔基体,并在高尔基体受蛋白酶加工剪切。加工成熟后的SREBP1进入细胞核,诱导胆固醇合成和脂质稳态维持相关基因的表达。进入细胞核的SREBP1会迅速地经历泛素化修饰,随后被蛋白酶降解。
SREBP1是多次跨膜蛋白,存在多种异构体,广泛表达于多种组织,在肝脏和肾上腺中表达丰度最高。
SREBP1的WB检测
SREBP1是一个多次跨膜蛋白,煮样可能导致蛋白聚集。如果煮样后检测不到信号,推荐提取蛋白时不煮样。
由于SREBP1会经历可变剪切,因此存在多种异构体,WB检测时有可能出现多条条带现象。
建议不要裁膜,或者保留40-250 kDa范围,防止蛋白信号丢失。
关键控制点
在实验中除了需要注意常规问题外,还要特别关注以下关键控制点:
样本制备
添加复合蛋白酶抑制剂以避免靶标蛋白降解。
选择合适的裂解液来富集更多靶标蛋白。
整个样本制备过程中,保持样本置于冰上。
通过Bradford分析、Lowry分析或BCA分析测定样本蛋白浓度。
电泳
至少上样20 μg总蛋白进行电泳。
建议使用阳性和阴性对照。
转膜
PVDF膜激活完成后充分清洗,完全去除膜上残留甲醇。
建议转膜完成后使用丽春红染色,确定转膜是否成功(如果选择荧光标记检测,请确保丽春红完全清洗干净)。
封闭
没有适用于所有体系的封闭液,请选择合适的封闭液。
1. L IJlst, H Mandel, W Oostheim, J P Ruiter, A Gutman, R J Wanders. Molecular basis of hepatic carnitine palmitoyltransferase I deficiency. Case Reports J Clin Invest. 1998 Aug 1;102(3):527-31. doi: 10.1172/JCI2927.
2. C Prip-Buus, L Thuillier, etc. Molecular and enzymatic characterization of a unique carnitine palmitoyltransferase 1A mutation in the Hutterite community. Mol Genet Metab. 2001 May;73(1):46-54. doi: 10.1006/mgme.2001.3176.
3. Stéphanie Gobin, Laure Thuillier, etc. Functional and structural basis of carnitine palmitoyltransferase 1A deficiency. J Biol Chem. 2003 Dec 12;278(50):50428-34. doi: 10.1074/jbc.M310130200. Epub 2003 Sep 29.
4. Mehmet Arif Icer, Makbule Gezmen-Karadag. The multiple functions and mechanisms of osteopontin. Clin Biochem. 2018 Sep;59:17-24. doi: 10.1016/j.clinbiochem.2018.07.003. Epub 2018 Jul 10.
5. Ashley Yim, Christian Smith, Amanda M Brown. Osteopontin/secreted phosphoprotein-1 harnesses glial-, immune-, and neuronal cell ligand-receptor interactions to sense and regulate acute and chronic neuroinflammation. Immunol Rev. 2022 Oct;311(1):224-233. doi: 10.1111/imr.13081. Epub 2022 Apr 22.
6. Anne-Sophie Lamort, Ioanna Giopanou, Ioannis Psallidas, Georgios T Stathopoulos. Osteopontin as a Link between Inflammation and Cancer: The Thorax in the Spotlight. Cells. 2019 Aug 2;8(8):815. doi: 10.3390/cells8080815.
7. C Yokoyama, X Wang, M R Briggs et al. SREBP-1, a basic-helix-loop-helix-leucine zipper protein that controls transcription of the low density lipoprotein receptor gene. Cell. 1993 Oct 8;75(1):187-97.
8. Y Hirano, M Yoshida, M Shimizu et al. Direct demonstration of rapid degradation of nuclear sterol regulatory element-binding proteins by the ubiquitin-proteasome pathway. J Biol Chem. 2001 Sep 28;276(39):36431-7. doi: 10.1074/jbc.M105200200.
9. Michiyo Amemiya-Kudo, Hitoshi Shimano, Alyssa H Hasty et al. Transcriptional activities of nuclear SREBP-1a, -1c, and -2 to different target promoters of lipogenic and cholesterogenic genes. J Lipid Res. 2002 Aug;43(8):1220-35.
Daqian Xu , Zheng Wang , Yan Xia et al. The gluconeogenic enzyme PCK1 phosphorylates INSIG1/2 for lipogenesis. Nature. 2020 Apr;580(7804):530-535. doi: 10.1038/s41586-020-2183-2.
